areps siddiqui etal 2013
TRANSCRIPT
Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/234145844
Psychrophiles
ArticleinAnnualReviewofEarthandPlanetarySciences·May2012
DOI:10.1146/annurev-earth-040610-133514
CITATIONS
13
READS
291
8authors,including:
KhawarSohailSiddiqui
KingFahdUniversityofPetroleumandMiner…
73PUBLICATIONS1,770CITATIONS
SEEPROFILE
TimothyWilliams
UNSWAustralia
63PUBLICATIONS1,581CITATIONS
SEEPROFILE
MichelleAllen
UNSWAustralia
22PUBLICATIONS613CITATIONS
SEEPROFILE
RicardoCavicchioli
UNSWAustralia
132PUBLICATIONS4,543CITATIONS
SEEPROFILE
Allin-textreferencesunderlinedinbluearelinkedtopublicationsonResearchGate,
lettingyouaccessandreadthemimmediately.
Availablefrom:KhawarSohailSiddiqui
Retrievedon:21August2016

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
RE V I E W
S
IN
AD V A
NC
E
PsychrophilesKhawar S. Siddiqui,1 Timothy J. Williams,1
David Wilkins,1 Sheree Yau,1 Michelle A. Allen,1
Mark V. Brown,1,2 Federico M. Lauro,1
and Ricardo Cavicchioli11School of Biotechnology and Biomolecular Sciences and 2Evolution and Ecology ResearchCenter, The University of New South Wales, Sydney, New South Wales 2052, Australia;email: [email protected]
Annu. Rev. Earth Planet. Sci. 2013. 41:6.1–6.29
The Annual Review of Earth and Planetary Sciences isonline at earth.annualreviews.org
This article’s doi:10.1146/annurev-earth-040610-133514
Copyright c© 2013 by Annual Reviews.All rights reserved
Keywords
microbial cold adaptation, cold-active enzymes, metagenomics, microbialdiversity, Antarctica
Abstract
Psychrophilic (cold-adapted) microorganisms make a major contributionto Earth’s biomass and perform critical roles in global biogeochemical cy-cles. The vast extent and environmental diversity of Earth’s cold biospherehas selected for equally diverse microbial assemblages that can include ar-chaea, bacteria, eucarya, and viruses. Underpinning the important ecologicalroles of psychrophiles are exquisite mechanisms of physiological adaptation.Evolution has also selected for cold-active traits at the level of molecularadaptation, and enzymes from psychrophiles are characterized by specificstructural, functional, and stability properties. These characteristics of en-zymes from psychrophiles not only manifest in efficient low-temperatureactivity, but also result in a flexible protein structure that enables biocatalysisin nonaqueous solvents. In this review, we examine the ecology of Antarcticpsychrophiles, physiological adaptation of psychrophiles, and properties ofcold-adapted proteins, and we provide a view of how these characteristicsinform studies of astrobiology.
6.1
Review in Advance first posted online on February 14, 2013. (Changes may still occur before final publication online and in print.)
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
INTRODUCTION
Much of life on Earth has evolved to colonize low-temperature environments. In fact, at tem-peratures permanently below 5◦C, the cold biosphere represents by far the largest fraction ofthe global biosphere (Feller & Gerday 2003, Cavicchioli 2006, Siddiqui & Cavicchioli 2006,Casanueva et al. 2010, Margesin & Miteva 2011). Consistent with representative size, the coldbiosphere consists of diverse types of environments—vast tracts of the deep sea, geographically dis-persed alpine regions, geologically specific subterranean caverns, climatically challenged regionsof permafrost, and biogeochemically diverse polar reaches (Figure 1). Proliferating throughoutthese cold realms is a plethora of psychrophilic (cold-adapted) microorganisms—archaea, bacte-ria, eucarya, and viruses. A small proportion of the isolated microorganisms from naturally coldenvironments have a restricted growth temperature range with an upper growth temperature limitless than ∼20◦C (stenopsychrophile), whereas the majority of isolates have a broader temperaturerange, tolerating warmer temperatures (eurypsychrophile).
Particularly through the application of molecular genetics approaches, most notably small sub-unit ribosomal RNA (SSU rRNA) sequencing, fluorescent in situ hybridization (FISH), and DNAsequencing of whole environmental samples (metagenomics), the cold biosphere has been discov-ered to harbor a diverse range of microbial groups. In recent years, the application of metagenomicsand associated meta-functional approaches (metaproteomics and metatranscriptomics) has shedlight on whole microbial community composition dynamics and microbial processes that are be-ing driven by the resident psychrophiles. Genomic, physiological, and biochemical analyses ofpsychrophilic isolates and their cellular components have also gleaned valuable information aboutthe diverse molecular mechanisms of cold adaptation. As a result, whether driven by global ques-tions concerning the impact of ecosystem change on microbial communities in cold environments,fundamental studies of molecular structure and function, or biotechnologically driven pursuits ofnovel cold-active biocatalysts, the field of psychrophiles has made great advances.
This review aims to cover topics relevant to studies of earth and planetary sciences by providingknowledge about physiological and protein adaptation—characteristics that speak to fundamentalprinciples of biological adaptation to the cold and provide insight into survivability. A perspectiveon microbial ecology of Antarctic systems opens the review, particularly focusing on lake, sea-ice,and deep-sea environments—systems that include a broad range of physicochemical conditions
Polar
e.g. Deep Lake,Antarctica
–20°C
Extraterrestrial
e.g. Europa
Surface: –200 to –160ºCSubsurface ocean: ?ºC
Alpine
< 10°C Deep sea
1 to 4°C
Figure 1Terrestrial and extraterrestrial cold environments. Representative temperatures are shown.
6.2 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
that provide knowledge about the diversity of microbial life that is sustained under a rangeof cold and abiotically varied environmental extremes. Also provided is a brief perspective onpsychrophiles and global warming, providing a glimpse into the use of cold-active enzymes andits impact on psychrophiles in relation to climate change. The review concludes with a sectionreflecting on microbial extremes and cold-active enzymes and their relevance to astrobiology.
ANTARCTIC PSYCHROPHILES
Antarctic Aquatic Ecosystems
Both southern and northern polar regions are delicately balanced ecosystems that are easily affectedby ecosystem changes (Moline et al. 2004, Murray & Grzymski 2007, Wilkins et al. 2012b), andglobal warming is expected to cause changes that will flow through to organisms right up thefood chain (Kirchman et al. 2009). In the Antarctic, global warming has particularly impactedthe Antarctic Peninsula and West Antarctica (Meredith & King 2005, Murray & Grzymski 2007,Cavalieri & Parkinson 2008, Whitehouse et al. 2008, Reid et al. 2009, Steig et al. 2009, Hogget al. 2011), and Antarctic sea-ice extent has decreased by at least ∼20% since the early 1950s andis projected to continue to decrease (Curran et al. 2003, Liu & Curry 2010). Ocean acidification(Kintisch & Stoksta 2008, McNeil & Matear 2008, Falkowski 2012), reduced CO2 absorption (LeQuere et al. 2007), and reduced nutrient supply particularly at higher latitudes caused by increasedstratification (Sarmiento & Le Quere 1996, Wignall & Twitchett 1996, Matear & Hirst 1999)are all effects linked to global warming. As the ocean microorganisms are critical for sequesteringanthropogenic CO2 (Sabine et al. 2004, Mikaloff Fletcher et al. 2006) and transporting it to thebenthic zones (Thomalla et al. 2011), the changes taking place in polar waters are of great concernfor the health of the global ecosystem.
Even though only 50,850 km2 (0.4%) of Antarctica is seasonally ice free (Poland et al. 2003,Cary et al. 2010), a broad range of lake systems are distributed around Antarctica that maintainice, water column, sediment, and microbial mat communities (Wilkins et al. 2012b). These lakesinclude subglacial, epiglacial, and surface systems that range in salinity from fresh to saturatedand from mixed to permanently stratified. The evolutionary history of these lakes is as varied asthe lakes themselves, which include the hundreds of marine-derived systems in the Vestfold Hills,which were isolated ∼3,000–7,000 years ago from the ocean (Gibson 1999) (Figure 2); subglacialoutflow from Blood Falls dating from 1.5 Mya (Mikucki et al. 2009); and waters in the depths ofsubglacial Lake Vostok, which are probably even older (Siegert et al. 2001).
Antarctic Microorganisms Colonize Diverse Cold Niches
Microbial populations vary in accordance with the wide range of physical and chemical propertiesof Antarctic lakes. In some marine-derived lakes, such as Ace Lake, the marine origin and, possibly,subsequent seeding from marine waters can be seen in the community composition of some partsof the water column (Lauro et al. 2011b) (Figure 2). However, this stratified system harbors vastlydifferent communities in other parts of the lake where very different physicochemical conditionsexist (Lauro et al. 2011b), including a highly purified population of green sulfur bacteria at the lake’soxycline interface (Ng et al. 2010). The microbial communities in Lake Bonney have evolved inresponse to physical distinctions occurring in two different lobes of the lake (Glatz et al. 2006). Bothof these examples illustrate how seed populations have diverged in response to ecosystem changes.
The transition from a marine to a hypersaline environment at Deep Lake provides an extremeexample of ecosystem change (Figure 2). Situated in the Vestfold Hills, Deep Lake is ∼55 m belowsea level, 36-m deep, hypersaline (3.6–4.8 M), ice free, and perennially cold (e.g., −20◦C) (Ferris &Burton 1988, Franzmann et al. 1988). The system appears on the border of sustaining life; scientific
www.annualreviews.org • Psychrophiles 6.3
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
a a b b
d d
a b c c c
f f
h h
f e
d
h
g
k
j i
m l n
6.4 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
records indicate it has been extremely unproductive (<10 g C m−2 year−1) (Campbell 1978).The microbial diversity in the lake is extremely low, dominated by members of the haloarchaea(Bowman et al. 2000a). Ongoing studies of this system have identified a range of genomic traitsand ecology of the system that are unique compared with hypersaline or cold aquatic systemselsewhere in the world (R. Cavicchioli, unpublished results).
Subsurface lake systems include subglacial lakes, such as Lake Vostok (Siegert et al. 2001), andepiglacial lakes that result from glacier melt and form where mountains (e.g., Framnes Moun-tains) penetrate the polar ice surface and may harbor microorganisms that are ancient or recent(postglacial) inhabitants (Gibson 2006, Cavicchioli 2007). Avoiding contamination in the pursuitof studying such pristine systems is a significant logistical challenge, and lessons learned aboutdrilling into Lake Vostok and other subglacial lakes (Inman 2005, Wingham et al. 2006, Alekhinaet al. 2007, Lukin & Bulat 2011, Gramling 2012, Jones 2012) should provide wisdom for guidingcontemplation of future endeavors, including extraterrestrial studies.
Antarctic Aquatic Microorganisms
Our understanding of community composition in Antarctic aquatic systems has been greatlyfacilitated by molecular-based studies (Wilkins et al. 2012b). These have included analyses usingdenaturing gradient gel electrophoresis (Pearce 2003, 2005; Pearce et al. 2003, 2005; Karr et al.2005; Unrein et al. 2005; Glatz et al. 2006; Mikucki & Priscu 2007; Mosier et al. 2007; Schiaffinoet al. 2009; Villaescusa et al. 2010), rRNA genes (Bowman et al. 2000a,b, 2003; Gordon et al.2000; Christner et al. 2001; Purdy et al. 2003; Karr et al. 2003, 2005, 2006; Matsuzaki et al. 2006;Kurosawa et al. 2010; Bielewicz et al. 2011), functional genes (Olsen et al. 1998, Voytek et al.1999, Mikucki et al. 2009), and metagenomics and metaproteomics (Lopez-Bueno et al. 2009; Nget al. 2010; Lauro et al. 2011b; Yau et al. 2011; Brown et al. 2012; Gryzmski et al. 2012; Varinet al. 2012; Wilkins et al. 2012a; Williams et al. 2012a,b).
Molecular signatures of archaea have been detected in a range of Antarctic lakes, includingstrictly anaerobic methanogens and aerobic haloarchaea (Bowman et al. 2000a,b; Purdy et al. 2003;
←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−Figure 2Antarctic lake systems. (a–k) Lakes in the Vestfold Hills (68◦33′0′ ′ S, 78◦15′0′ ′ E) and (l–n) Heard Island (53◦6′0′ ′ S, 73◦31′0′ ′E).(a) Ace Lake, a marine-derived meromictic system (Gibson 1999, Cavicchioli 2006) that is separated from marine waters of Long Fjordby only several hundred meters ( foreground ). Sea ice and icebergs are present in the early-mid austral summer 2008 (background ).Among other species, green sulfur bacteria play a particularly important role in this lake’s ecosystem (Ng et al. 2010, Lauro et al.2011b). (b) Ace Lake at the end of summer 2006 after the lake ice and sea ice have melted and begun to refreeze. (c) Snow drifts on AceLake formed after a blizzard behind quad bikes (used for transport between the lake and Davis Research Base located 15 km away) andmobile work shelters (MWSs) that were used for sample collection and protection from the weather. (d ) Organic Lake, a hypersalinemeromictic system where the waters are −13◦C below the surface ice; photo taken in 2008 (Gibson 1999). The novel and importantrole of virophages was discovered in this lake (Yau et al. 2011). (e) Drilling through surface ice on Organic Lake prior to the positioningof MWSs for sample collection. ( f ) Foam generated by the wind blowing across the organically rich waters of Organic Lake in 2006.Shown are microbial biofilms (orange) in the water and on rocks as well as penguin feathers (white) near the edge of the lake. ( g) DeepLake panorama in September 2008 after a cold winter (−40◦C) (photo credit: Mark Milnes). (h) Deep Lake is hypersaline, and watertemperatures reach −20◦C and do not freeze. (i ) Deep Lake is ∼55 m below sea level, marked by the flat hill line in the background.( j ) The Vestfold Hills region contains hundreds of lakes and ponds positioned between the coastline and the edge of the Antarcticcontinental ice mass (background ). (k) MWSs, dinghy, and research equipment at Deep Lake. Water pumped into drums on board thedinghy at the center of the lake (∼800 m from shore) was transported back to the MWSs for processing. (l ) Brown Lagoon at the baseof Brown Glacier, Heard Island, in 2008 contains glacier meltwater and is separated from ocean waters by a narrow strip of beach.(m) Winston Lagoon at the base of Winston Glacier is open to the ocean, allowing water exchange. (n) Water formed at the base ofStephenson Glacier contains slabs of recently melted glacier. The melted sections and large lake of water that were not present inprevious seasons are overt signs of ecosystem change as a result of global warming.
www.annualreviews.org • Psychrophiles 6.5
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Glatz et al. 2006; Karr et al. 2006; Kurasawa et al. 2010; Lauro et al. 2011b), of which several havebeen brought into axenic culture (Franzmann et al. 1988, 1992, 1997). A large number of studieshave focused on Antarctic bacteria, and diverse taxa have been identified, including members ofthe groups Actinobacteria, Alphaproteobacteria, Betaproteobacteria, Chlorobi, Chloroflexi, Cyanobacteria,Deltaproteobacteria, Firmicutes, Gammaproteobacteria, Bacteroidetes [Cytophaga-Flavobacterium-Bacteroides (CFB) group], Planctomycetes, Spirochaetes, and Verrucomicrobia (Bowman et al. 2000a,b;Glatz et al. 2006; Mosier et al. 2007; Kurosawa et al. 2010; Pearce 2005; Pearce et al. 2003,2005; Schiaffino et al. 2009; Lauro et al. 2011b). Eucarya, particularly algal phototrophs, are alsoimportant in Antarctic lakes, although fungi and silicoflagellates have also been identified (Unreinet al. 2005, Mosier et al. 2007, Bielewicz et al. 2011, Lauro et al. 2011b, Yau et al. 2011). Virusesof Antarctic eucarya, bacteria, and archaea have also been identified (Lauro et al. 2011b, Yau et al.2011). The absence of higher trophic level organisms in Antarctic lake systems indicates virusesmay play an important role in the microbial loop (Kepner et al. 1998; Anesio & Bellas 2011;Laybourn-Parry et al. 2001, 2007; Madan et al. 2005; Sawstrom et al. 2007; Lopez-Bueno et al.2009). Specific impacts on bacterial hosts have been linked to mechanisms of cellular resistance;uncharacteristically low levels of viruses (Lauro et al. 2011b); and roles for virophage predationof algal viruses, which is predicted to increase overall primary production and net carbon flow inthe lake system (Yau et al. 2011) (Figure 2).
In addition to the water column, rich microbial communities are found in Antarctic mats andcan make important contributions to biomass and productivity (Vincent 2000, Moorhead et al.2005, Laybourn-Parry & Pearce 2007). Microorganisms identified in Antarctic mats includemembers of Actinobacteria, CFB, Cyanobacteria, Deinococcus-Thermus, Firmicutes, fungi, greenalgae, Planctomycetes, Proteobacteria, and Verrucomicrobia (Brambilla et al. 2001; Van Trappen et al.2002; Taton et al. 2003, 2006; Jungblut et al. 2005; Fernandez-Valiente et al. 2007; Sutherland2009; Borghini et al. 2010; Verleyen et al. 2010; Anderson et al. 2011; Callejas et al. 2011;Fernandez-Carazo et al. 2011; Hawes et al. 2011; Peeters et al. 2011, 2012; Antibus et al. 2012a,b;Varin et al. 2012). Mats are interesting features of lakes because they provide mineral andbiological records of the ecosystem, thereby also providing insight into the evolution of past andextant species (Bomblies et al. 2001, Sutherland & Hawes 2009, Anderson et al. 2011, Hawes et al.2011).
The taxa in Antarctic marine waters are, on the whole, similar to those in temperate or tropicalocean waters and include a high proportion of Alphaproteobacteria (e.g., SAR11 clade), Flavobacteria,Gammaproteobacteria, and ammonia oxidizing Marine Group I Crenarchaeota (Wilkins et al. 2012b).However, although many common taxa are found, the indigenous Antarctic populations havegenetic and physiological traits that enable them to compete effectively at low temperatures andunder the specific physicochemical regimes that prevail (e.g., Brown et al. 2012).
Molecular analyses offer insight into microbial communities because they can canvass largecross sections of the community (e.g., pyrotag sequencing of SSU rRNA genes) and particularlybecause they report on the whole community irrespective of whether the microorganisms areamenable to cultivation—the majority of which are not (Amann et al. 1995). However, althoughmolecular analyses have proven useful for studying sea-ice microorganisms, a high proportionof these communities are culturable and, hence, amenable to laboratory study. Antarctic isolatesinclude members of the genera Arthrobacter, Colwellia, Gelidibacter, Glaciecola, Halobacillus,Halomonas, Hyphomonas, Marinobacter, Planococcus, Pseudoalteromonas, Pseudomonas, Psychrobacter,Psychroflexus, Psychroserpens, Shewanella, and Sphingomonas (Bowman et al. 1997a–c, 1998a,b).Sea-ice communities have adapted to a range of location-specific physicochemical conditions,including temperature (0 to −35◦C), salinity (up to seven times seawater salinity), pH, light, andnutrient gradients (Eicken 2003, Mock & Thomas 2005).
6.6 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
PHYSIOLOGICAL ADAPTATIONS IN PSYCHROPHILES
Overview
Physiological adaptations to growth temperature can be identified by comparing the propertiesof microorganisms that grow naturally at different temperatures. However, compared with pro-tein adaptation (see below) where insight can be gained by comparing the properties of proteinsbetween psychrophiles and hyper/thermophiles, physiological adaptation is more complicatedowing to the greater number of factors that can impact the complex variety of components in acell and ultimately cause an adaptive response. The cell’s physiology is dictated by its genomiccomplement of genes and the regulation of gene expression in response to environmental stimuli.Depending on the environment, a large number of biotic (e.g., predation by grazers and viruses,antibiotics, cell-cell interactions), abiotic (e.g., pH, salinity, oxygen, nutrient flux), and broaderecological factors (e.g., sea ice versus seawater, particle attached versus free living) can greatlyinfluence the selection and growth properties of individual microorganisms. In addition, the di-versity of microorganisms colonizing Earth’s biosphere, the majority of which is cold, is enormous.As a result, a variety of physiotypes have evolved to colonize cold environments successfully. Inaddition, very few classes of microorganisms that can successfully colonize both low- and high-temperature extremes have evolved. Methanogens, which are members of Archaea, are the onlygroup known to have individual species that span the growth temperature range from subzero to122◦C (Saunders et al. 2003, Cavicchioli 2006, Reid et al. 2006, Takai et al. 2008). Thus, thereare limited opportunities to compare the adaptive traits of psychrophiles and hyper/thermophilesthat belong to the same genus or family.
As a result, most of our knowledge about physiological adaptations has been gained by ex-amining the response of individual microorganisms to different growth temperatures (e.g., highversus low temperature). In this respect, global expression studies (e.g., proteomics, transcrip-tomics) linked to knowledge of direct physiological measurements (e.g., temperature and nutrientperturbation of morphology, growth rate, rates of macromolecular synthesis, solute composition,membrane lipid composition, modification of nucleic acids) have proven particularly valuablefor determining the mechanisms of psychrophile adaptation (see, for example, Cavicchioli 2006).Examples of knowledge gained are described below.
Cellular Mechanisms of Cold Adaptation
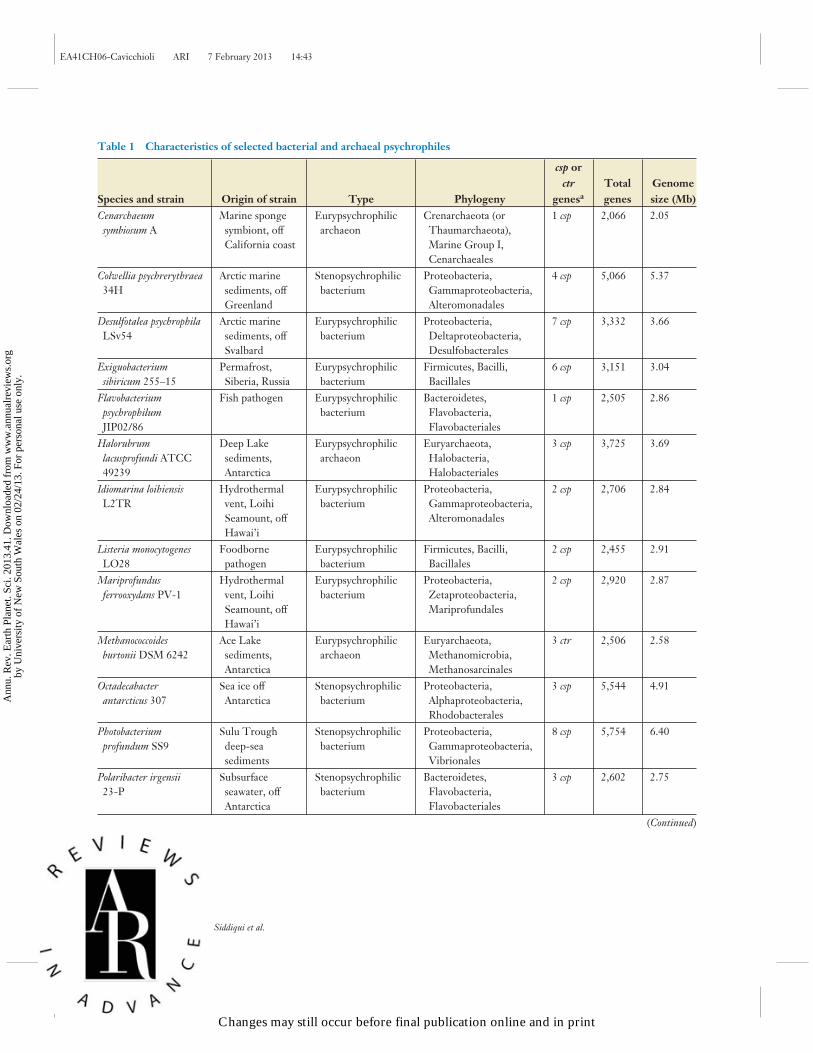
Low temperature can impede transcription and translation owing to the increased stability ofadventitious secondary structures of transcripts. Preventing or resolving inhibitory secondarystructures of RNA can be achieved by RNA chaperones. Cold shock proteins (Csps) are smallproteins that bind to RNA to preserve its single-stranded conformation ( Jones & Inouye 1994).DEAD box RNA helicases are capable of unwinding secondary structures in an ATP-dependentmanner and are upregulated during cold growth in some psychrophiles (Lim et al. 2000). Psy-chrophiles vary widely in the number of csp genes present in their genomes (Table 1). Csps containa nucleic-acid-binding domain, known as the cold shock domain (CSD), and have additional rolesbesides serving as RNA chaperones. Individual CSD-containing proteins can regulate the coldshock response or play a major role in subsequent growth at low temperatures in mesophiles(Hebraud & Potier 1999). Thus, many of the Csps act as cold-adaptive proteins in psychrophiles,because they are constitutively rather than transiently expressed at low temperatures (D’Amicoet al. 2006). Overexpression of cspA of Psychromonas arctica was shown to increase cold resistanceof Escherichia coli at low temperatures ( Jung et al. 2010). Additionally, one of three Csps appearsto be important in the low-temperature growth of Shewanella oneidensis (Gao et al. 2006).
www.annualreviews.org • Psychrophiles 6.7
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.
EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Table 1 Characteristics of selected bacterial and archaeal psychrophiles
Species and strain Origin of strain Type Phylogeny
csp orctr
genesaTotalgenes
Genomesize (Mb)
Cenarchaeumsymbiosum A
Marine spongesymbiont, offCalifornia coast
Eurypsychrophilicarchaeon
Crenarchaeota (orThaumarchaeota),Marine Group I,Cenarchaeales
1 csp 2,066 2.05
Colwellia psychrerythraea34H
Arctic marinesediments, offGreenland
Stenopsychrophilicbacterium
Proteobacteria,Gammaproteobacteria,Alteromonadales
4 csp 5,066 5.37
Desulfotalea psychrophilaLSv54
Arctic marinesediments, offSvalbard
Eurypsychrophilicbacterium
Proteobacteria,Deltaproteobacteria,Desulfobacterales
7 csp 3,332 3.66
Exiguobacteriumsibiricum 255–15
Permafrost,Siberia, Russia
Eurypsychrophilicbacterium
Firmicutes, Bacilli,Bacillales
6 csp 3,151 3.04
FlavobacteriumpsychrophilumJIP02/86
Fish pathogen Eurypsychrophilicbacterium
Bacteroidetes,Flavobacteria,Flavobacteriales
1 csp 2,505 2.86
Halorubrumlacusprofundi ATCC49239
Deep Lakesediments,Antarctica
Eurypsychrophilicarchaeon
Euryarchaeota,Halobacteria,Halobacteriales
3 csp 3,725 3.69
Idiomarina loihiensisL2TR
Hydrothermalvent, LoihiSeamount, offHawai’i
Eurypsychrophilicbacterium
Proteobacteria,Gammaproteobacteria,Alteromonadales
2 csp 2,706 2.84
Listeria monocytogenesLO28
Foodbornepathogen
Eurypsychrophilicbacterium
Firmicutes, Bacilli,Bacillales
2 csp 2,455 2.91
Mariprofundusferrooxydans PV-1
Hydrothermalvent, LoihiSeamount, offHawai’i
Eurypsychrophilicbacterium
Proteobacteria,Zetaproteobacteria,Mariprofundales
2 csp 2,920 2.87
Methanococcoidesburtonii DSM 6242
Ace Lakesediments,Antarctica
Eurypsychrophilicarchaeon
Euryarchaeota,Methanomicrobia,Methanosarcinales
3 ctr 2,506 2.58
Octadecabacterantarcticus 307
Sea ice offAntarctica
Stenopsychrophilicbacterium
Proteobacteria,Alphaproteobacteria,Rhodobacterales
3 csp 5,544 4.91
Photobacteriumprofundum SS9
Sulu Troughdeep-seasediments
Stenopsychrophilicbacterium
Proteobacteria,Gammaproteobacteria,Vibrionales
8 csp 5,754 6.40
Polaribacter irgensii23-P
Subsurfaceseawater, offAntarctica
Stenopsychrophilicbacterium
Bacteroidetes,Flavobacteria,Flavobacteriales
3 csp 2,602 2.75
(Continued)
6.8 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.
EA41CH06-Cavicchioli ARI 7 February 2013 14:43
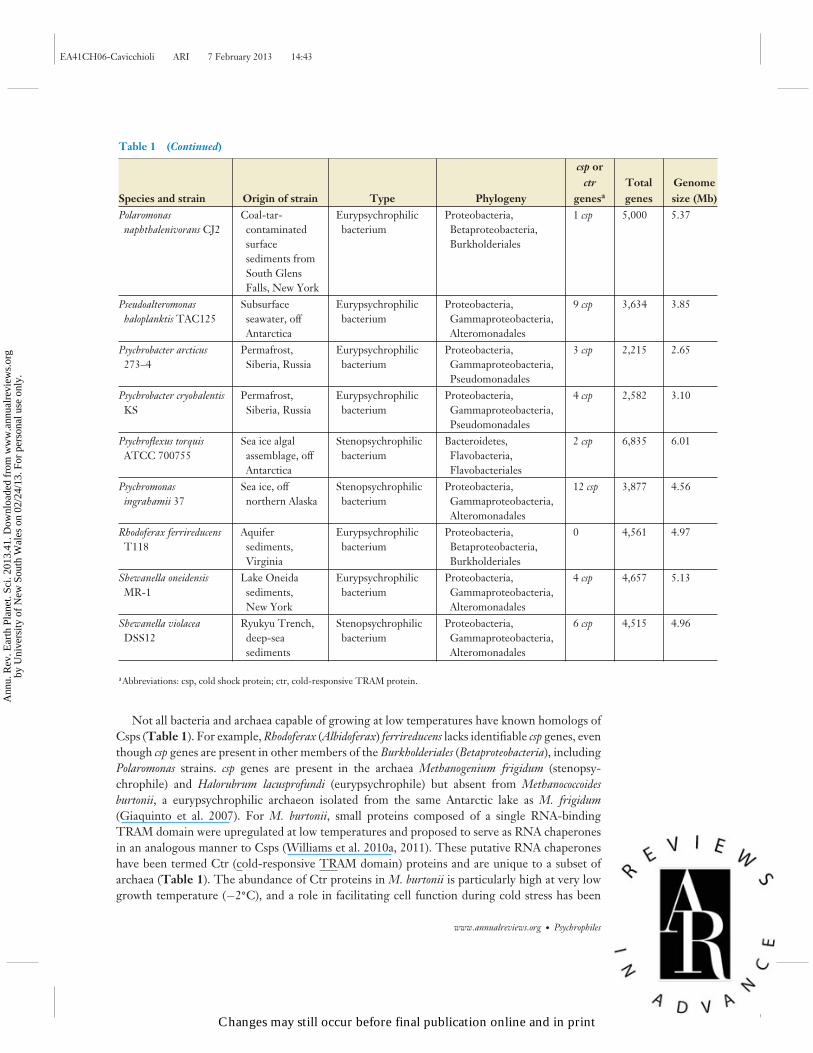
Table 1 (Continued)
Species and strain Origin of strain Type Phylogeny
csp orctr
genesaTotalgenes
Genomesize (Mb)
Polaromonasnaphthalenivorans CJ2
Coal-tar-contaminatedsurfacesediments fromSouth GlensFalls, New York
Eurypsychrophilicbacterium
Proteobacteria,Betaproteobacteria,Burkholderiales
1 csp 5,000 5.37
Pseudoalteromonashaloplanktis TAC125
Subsurfaceseawater, offAntarctica
Eurypsychrophilicbacterium
Proteobacteria,Gammaproteobacteria,Alteromonadales
9 csp 3,634 3.85
Psychrobacter arcticus273–4
Permafrost,Siberia, Russia
Eurypsychrophilicbacterium
Proteobacteria,Gammaproteobacteria,Pseudomonadales
3 csp 2,215 2.65
Psychrobacter cryohalentisKS
Permafrost,Siberia, Russia
Eurypsychrophilicbacterium
Proteobacteria,Gammaproteobacteria,Pseudomonadales
4 csp 2,582 3.10
Psychroflexus torquisATCC 700755
Sea ice algalassemblage, offAntarctica
Stenopsychrophilicbacterium
Bacteroidetes,Flavobacteria,Flavobacteriales
2 csp 6,835 6.01
Psychromonasingrahamii 37
Sea ice, offnorthern Alaska
Stenopsychrophilicbacterium
Proteobacteria,Gammaproteobacteria,Alteromonadales
12 csp 3,877 4.56
Rhodoferax ferrireducensT118
Aquifersediments,Virginia
Eurypsychrophilicbacterium
Proteobacteria,Betaproteobacteria,Burkholderiales
0 4,561 4.97
Shewanella oneidensisMR-1
Lake Oneidasediments,New York
Eurypsychrophilicbacterium
Proteobacteria,Gammaproteobacteria,Alteromonadales
4 csp 4,657 5.13
Shewanella violaceaDSS12
Ryukyu Trench,deep-seasediments
Stenopsychrophilicbacterium
Proteobacteria,Gammaproteobacteria,Alteromonadales
6 csp 4,515 4.96
aAbbreviations: csp, cold shock protein; ctr, cold-responsive TRAM protein.
Not all bacteria and archaea capable of growing at low temperatures have known homologs ofCsps (Table 1). For example, Rhodoferax (Albidoferax) ferrireducens lacks identifiable csp genes, eventhough csp genes are present in other members of the Burkholderiales (Betaproteobacteria), includingPolaromonas strains. csp genes are present in the archaea Methanogenium frigidum (stenopsy-chrophile) and Halorubrum lacusprofundi (eurypsychrophile) but absent from Methanococcoidesburtonii, a eurypsychrophilic archaeon isolated from the same Antarctic lake as M. frigidum(Giaquinto et al. 2007). For M. burtonii, small proteins composed of a single RNA-bindingTRAM domain were upregulated at low temperatures and proposed to serve as RNA chaperonesin an analogous manner to Csps (Williams et al. 2010a, 2011). These putative RNA chaperoneshave been termed Ctr (cold-responsive TRAM domain) proteins and are unique to a subset ofarchaea (Table 1). The abundance of Ctr proteins in M. burtonii is particularly high at very lowgrowth temperature (−2◦C), and a role in facilitating cell function during cold stress has been
www.annualreviews.org • Psychrophiles 6.9
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
proposed (Williams et al. 2011). The upregulation of Ctr proteins in M. burtonii in response togrowth in the presence of the solvent methanol further suggests a wider role in the cell as stressresponse proteins (Williams et al. 2010a).
Small RNA-binding proteins (Rbps) can facilitate cold adaptation, but similar to Csps, theycan also have other functional roles in the cell (Maruyama et al. 1999, Christiansen et al. 2004).These Rbps accumulate following cold stress and play important roles in regulating transcriptiontermination (Mori et al. 2003), Rbps are small proteins that contain a single glycine-rich RNA-binding motif. They are prevalent in cyanobacteria but rare in other bacteria (Maruyama et al.1999, Ehira et al. 2003). The mesophilic cyanobacterium Anabaena variabilis has eight rbp genes,all but one of which are cold regulated (Maruyama et al. 1999). Osmotic stress also enhances rbpgene expression in Anabaena sp. PCC 7120: Responses to cold and osmotic stresses overlap becausethey both decrease the availability of free water (Mori et al. 2003). Rbp proteins may also play arole in thermal adaptation in psychrophilic cyanobacteria, as expression of rbp genes increases atlow temperatures in the Antarctic strain Oscillatoria sp. SU1 (Ehira et al. 2003).
Nucleoside modifications can affect the stability of tRNA. As a result, the extent of modificationtends to be high in hyperthermophilic archaea and bacteria (Dalluge et al. 1997, Noon et al. 2003).However, dihydrouridine can enhance tRNA flexibility and is elevated in some psychrophilicbacteria and archaea (Dalluge et al. 1997, Noon et al. 2003).
Enzymes involved in the degradation of RNA and proteins are upregulated during low-temperature growth in some psychrophilic bacteria and archaea, including RNases and pro-teases from the permafrost bacterium Psychrobacter arcticus (Bergholz et al. 2009) and M. burtonii(Williams et al. 2010b). This has been interpreted as a strategy to conserve biosynthetic precursors(Bergholz et al. 2009) or as enhanced quality control of irreparably damaged RNA and proteins(Williams et al. 2010b), although the two are not mutually exclusive.
Energy conservation and biosynthetic pathways can be regulated in response to low-temperature growth. Psychrobacter cryohalolentis, a eurypsychrophilic bacterium isolated fromSiberian permafrost, increases the cytoplasmic pool of ATP and ADP to offset reduced ATP-dependent reaction rates (Amato & Christner 2009). Specific carbon substrate utilization pathways(e.g., methanol versus trimethylamine) are differentially regulated with growth temperature in M.burtonii (Williams et al. 2010a,b). In P. arcticus, a large number of energy metabolism genes aredownregulated at low temperatures (Bergholz et al. 2009), whereas P. cryohalolentis shows upreg-ulation of glyoxylate cycle enzymes (Bakermans et al. 2007). These examples highlight the varietyand complexity of metabolic responses of individual psychrophiles.
At temperatures low enough for ice to form, cells are subjected to additional stressors such asice damage, oxidative insult, and osmotic imbalance (Tanghe et al. 2003; Williams et al. 2010b,2011). Extracellular polymeric substances (EPS) can offer protection against mechanical disruptionto the cell membrane caused by ice. Sea-ice bacteria such as Colwellia psychrerythraea producepolysaccharide-rich EPS (Thomas & Dieckmann 2002, Junge et al. 2004). The resulting biofilmsmay afford protection against invasive ice crystal damage as well as facilitate the acquisition ofnutrients within the channels that form within the sea ice (Thomas & Dieckmann 2002; Jungeet al. 2004; Mancuso Nichols et al. 2005a,b). At low temperatures, psychrophilic archaea such as H.lacusprofundi and M. burtonii also form multicellular aggregates embedded in EPS (Reid et al. 2006).
Low temperatures decrease membrane fluidity and permeability. In response, the elastic liq-uid crystalline nature of the cell membrane is replaced by a gel-phase state that can impair thebiological functions of the membrane, including transport (Phadtare 2004). This can be offset byincreasing the proportion of unsaturated fatty acids in the lipid bilayer, resulting in a more looselypacked array (Russell 2008). Increasing the proportion of unsaturated fatty acids can be achievedby decreasing the saturation of pre-existing fatty acids or by synthesizing fewer saturated fatty acids
6.10 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
de novo. The eurypsychrophilic bacterium Exiguobacterium sibiricum has higher fatty acid desat-urase gene expression at low temperatures (Ponder et al. 2005, Rodrigues et al. 2008). M. burtonii,which lacks a fatty acid desaturase, alters expression of several lipid biosynthesis genes, resultingin fewer saturated isoprenoid lipid precursors (Nichols et al. 2004). Unsaturated isoprenoid lipidshave also been detected in H. lacusprofundi (Gibson et al. 2005). Many psychrophilic membersof Gammaproteobacteria (e.g., species of Colwellia, Moritella, Photobacterium, Psychromonas, Mari-nomonas, and Shewanella) are characterized by a high proportion of unsaturated fatty acids in theircell membranes (Margesin & Miteva 2011). In a metagenomic analysis, a microbial assemblage inglacier ice was found to be relatively enriched for genes involved in the maintenance of membranefluidity (Simon et al. 2009). Membrane lipid changes appear to be a generally conserved featurefor cellular adaptation to the cold.
Adaptation of Psychrophiles Viewed Through Genomes and Global GeneExpression Profiles
Many of the advances in understanding adaptive mechanisms have come from studies involvingthe genome sequences of psychrophiles. Approximately 30 bacterial and 4 archaeal genomesequences are available for psychrophiles originating from diverse cold habitats that includeAntarctic lakes, symbionts of sea sponges, marine sediment, permafrost, marshes, fish pathogens,and Kimchi (Lauro et al. 2011a). In addition to providing genomic blueprints that describethe capacity of psychrophiles, genomes provide the basis for targeted and global functionalstudies (e.g., proteomics and transcriptomics). The capacity to overview global responses isgreatly accelerating the ways in which knowledge is being gained about adaptive mechanisms, inparticular, as researchers define general characteristics of psychrophilic microorganisms versusspecific traits of individual psychrophiles.
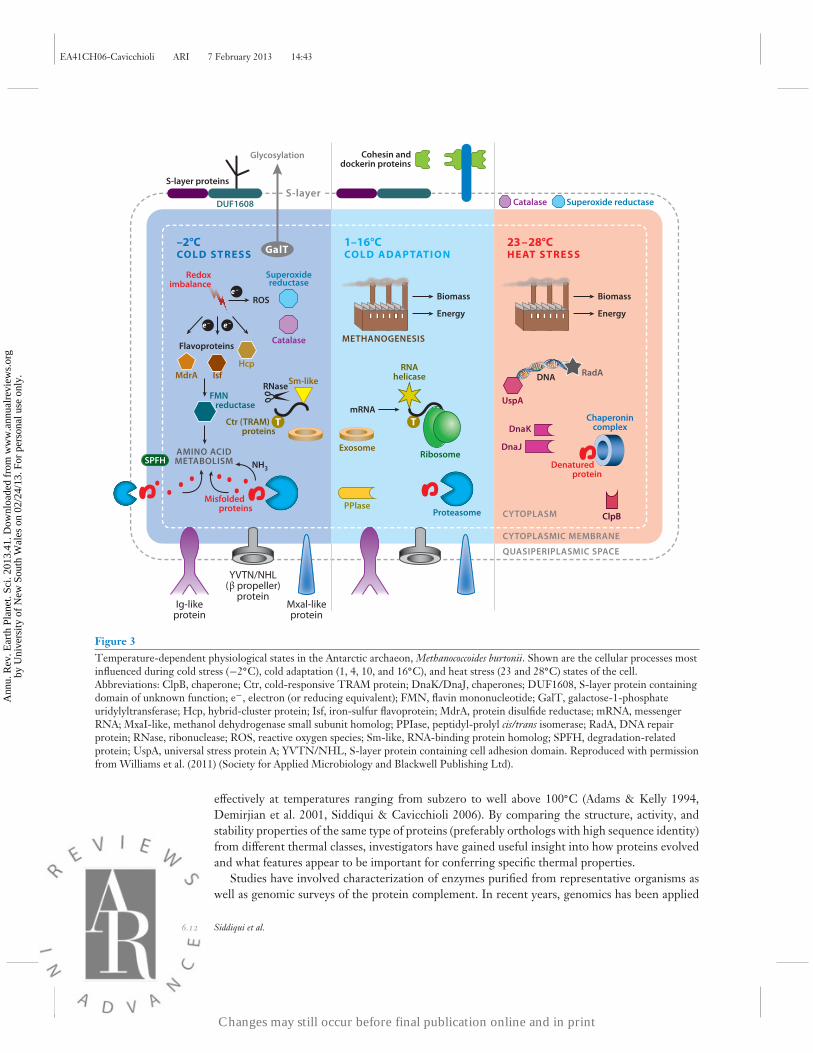
Good illustrations of what can be defined by these approaches include recent analyses of expres-sion profiles across multiple growth temperatures. An analysis of P. arcticus (growth temperaturerange from −10◦C to 28◦C) used transcriptomics to identify differences in mRNA abundancebetween four growth temperatures (−6, 0, 17, and 22◦C) (Bergholz et al. 2009), and a multiplexproteomics study of M. burtonii quantitated changes occurring across seven growth temperaturesthat span the organism’s complete growth temperature range (−2◦C to 28◦C) (Williams et al.2011) (Figure 3). In the latter study, by including growth temperature extremes as well as tem-peratures in between, researchers were able to infer stressful versus nonstressful physiologicalstates. Interestingly, the upregulation of oxidative stress proteins at both upper and lower tem-perature extremes demonstrated the important, yet distinct, ways in which temperature-inducedoxidative stress manifests in the cell. The study also revealed that protein profiles at temperaturesin which M. burtonii grew fastest (Topt) were similar to those at maximum growth temperature(Tmax). These findings highlighted the extent to which this psychrophile was heat stressed at thesetemperatures, which is consistent with a number of other studies that suggest that psychrophilesgrowing at Topt are likely to be heat stressed (Feller & Gerday 2003; Bakermans & Nealson 2004;Goodchild et al. 2004; Cavicchioli 2006; Williams et al. 2010b, 2011).
PROTEIN ADAPTATION TO THE COLD
Overview
Many types of proteins, including diverse classes of enzymes (e.g., glucanases, hydrolases, oxidore-ductases, hydrogenases, isomerases, nucleic acid-modifying enzymes), have evolved to function
www.annualreviews.org • Psychrophiles 6.11
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.
EA41CH06-Cavicchioli ARI 7 February 2013 14:43
e–
e–e–
TT
GalT
Glycosylation
Ig-likeprotein
Mxal-likeprotein
YVTN/NHL(β propeller)
protein
METHANOGENESIS
AMINO ACIDMETABOLISM
Flavoproteins
Biomass
Energy
Biomass
Energy
MdrA Isf
Hcp
Sm-like
ClpB
DnaJ
DnaK
Catalase
CatalaseDUF1608
UspA
RadA
FMN reductase
Proteasome
Exosome
Ctr (TRAM) proteins
RNAhelicase
PPlase
S-layer proteins
Cohesin anddockerin proteins
Ribosome
Chaperonincomplex
Misfolded proteins
Denatured protein
S-layerS-layerS-layer
CYTOPLASM
CYTOPLASMIC MEMBRANE
QUASIPERIPLASMIC SPACE
Superoxide reductase
Superoxidereductase
ROS
mRNA
RNaseDNA
NH3
Redoximbalance
–2°CCOLD STRESS
1–16°CCOLD ADAPTATION
23–28°CHEAT STRESS
SPFH
Figure 3Temperature-dependent physiological states in the Antarctic archaeon, Methanococcoides burtonii. Shown are the cellular processes mostinfluenced during cold stress (−2◦C), cold adaptation (1, 4, 10, and 16◦C), and heat stress (23 and 28◦C) states of the cell.Abbreviations: ClpB, chaperone; Ctr, cold-responsive TRAM protein; DnaK/DnaJ, chaperones; DUF1608, S-layer protein containingdomain of unknown function; e−, electron (or reducing equivalent); FMN, flavin mononucleotide; GalT, galactose-1-phosphateuridylyltransferase; Hcp, hybrid-cluster protein; Isf, iron-sulfur flavoprotein; MdrA, protein disulfide reductase; mRNA, messengerRNA; MxaI-like, methanol dehydrogenase small subunit homolog; PPIase, peptidyl-prolyl cis/trans isomerase; RadA, DNA repairprotein; RNase, ribonuclease; ROS, reactive oxygen species; Sm-like, RNA-binding protein homolog; SPFH, degradation-relatedprotein; UspA, universal stress protein A; YVTN/NHL, S-layer protein containing cell adhesion domain. Reproduced with permissionfrom Williams et al. (2011) (Society for Applied Microbiology and Blackwell Publishing Ltd).
effectively at temperatures ranging from subzero to well above 100◦C (Adams & Kelly 1994,Demirjian et al. 2001, Siddiqui & Cavicchioli 2006). By comparing the structure, activity, andstability properties of the same type of proteins (preferably orthologs with high sequence identity)from different thermal classes, investigators have gained useful insight into how proteins evolvedand what features appear to be important for conferring specific thermal properties.
Studies have involved characterization of enzymes purified from representative organisms aswell as genomic surveys of the protein complement. In recent years, genomics has been applied
6.12 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
to microbial communities from whole environmental samples (metagenomics), thereby providingDNA sequence information for proteins from uncultivated microorganisms. Metagenomics ofsamples from cold environments has included the generation of large data sets obtained by shotgunsequencing (e.g., Lopez-Bueno et al. 2009, Lauro et al. 2011b, Yau et al. 2011, Brown et al. 2012,Varin et al. 2012, Wilkins et al. 2012a, Williams et al. 2012b) and functional screening of clonesfor cold-active enzymes (e.g., Elenda et al. 2007, Kim et al. 2009). Genomic and metagenomicanalyses facilitate subsequent targeted analyses to assess specific features of individual proteins (e.g.,site-directed mutagenesis). Broad-spectrum modification (e.g., mutagenesis by directed evolution,chemical modification of particular amino acid side groups) and assessment of changes in thermalproperties of individual enzymes have also been used to identify structural properties that play rolesin conferring thermal activity/stability (Cavicchioli et al. 2006, Siddiqui et al. 2006). Collectively,these types of studies have revealed a great deal about the adaptation of proteins to temperature.
To achieve sufficient structural flexibility to afford enzyme activity at low temperatures, en-zymes have evolved specific compositional biases (i.e., amino acid composition) and secondary,tertiary, and/or quaternary structural properties (Feller & Gerday 2003, D’Amico et al. 2006,Siddiqui & Cavicchioli 2006, Feller 2008). In contrast, proteins from hyper/thermophiles requiresufficient structural rigidity to resist unfolding, which is also manifested through specific com-positional and structural properties (Daniel et al. 2008). In general terms, the features associatedwith adaptation (e.g., proportion of specific amino acids, hydrophobicity of exposed surfaces) tendto have opposite trends between proteins from psychrophiles and those from hyper/thermophiles(Siddiqui & Cavicchioli 2006, Feller 2008).
Proteins from psychrophiles have higher activity and thermolability compared with mesophilicand thermophilic homologues (Demirjian et al. 2001, Siddiqui & Cavicchioli 2006). For exam-ple, α-amylases from the psychrophilic bacterium Pseudoalteromonas haloplanktis and from thethermophilic bacterium Bacillus amyloliquefaciens have an optimal temperature of activity (Topt)of 28◦C and 84◦C, respectively (D’Amico et al. 2003). A striking example of cold adaptation isalanine racemase from Bacillus psychrosaccharolyticus, which has a Topt of 0◦C (Okubo et al. 1999). Be-cause low-temperature environments present significant problems for enzyme and, more broadly,protein function, the unique properties of cold-active enzymes has attracted both academic andcommercial interest (Cavicchioli et al. 2002, Feller & Gerday 2003, Cavicchioli & Siddiqui 2006,Siddiqui & Cavicchioli 2006, Feller 2008, Cavicchioli et al. 2011). This has led to rapid growthin the description of enzymes from a broad range of psychrophiles, with a concomitant devel-opment of biochemical and biophysical approaches attuned to their characterization (Feller &Gerday 2003, Cavicchioli et al. 2006, Siddiqui & Cavicchioli 2006). Below we discuss some of themechanisms by which thermal adaptation at low temperatures is attained.
Mechanisms of Enzyme Adaptation to the Cold
In low-temperature environments, there is insufficient kinetic energy to overcome enzyme acti-vation barriers, thus resulting in very slow rates of chemical reactions. For a biochemical reactionoccurring in a mesophile at 37◦C, a drop in temperature from 37◦C to 0◦C results in a 20–80-foldreduction in enzyme activity. This is the main factor preventing growth at low temperatures.However, organisms adapted to low temperatures have evolved several ways to overcome thisconstraint, including the energetically costly strategy of enhanced enzyme production (Crawford& Powers 1992) and seasonal expression of isoenzymes (Somero 1995). However, the most com-mon adaptive feature of cold-active enzymes is a reaction rate (kcat) that is largely independentof temperature. The majority of psychrophilic enzymes achieve temperature-insensitive kcat bydecreasing the activation energy barrier between the ground state (substrate) and activated state
www.annualreviews.org • Psychrophiles 6.13
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.
EA41CH06-Cavicchioli ARI 7 February 2013 14:43
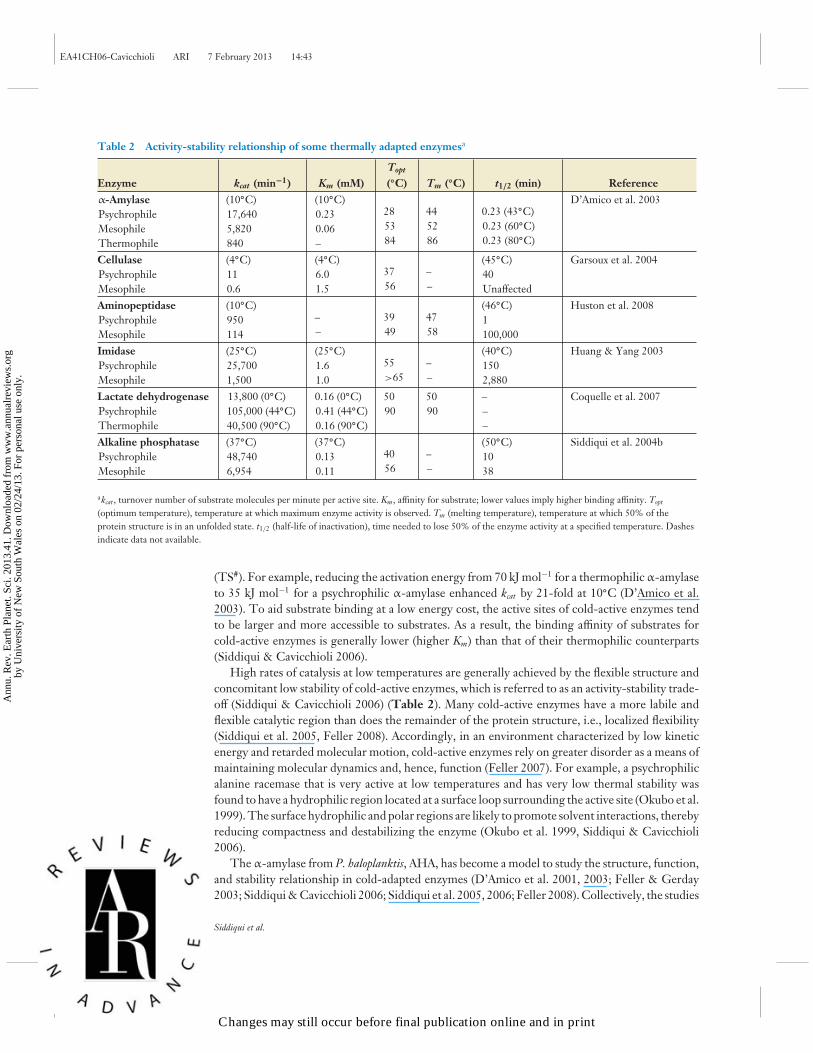
Table 2 Activity-stability relationship of some thermally adapted enzymesa
Enzyme kcat (min−1) Km (mM)Topt
(◦C) Tm (◦C) t1/2 (min) Referenceα-AmylasePsychrophileMesophileThermophile
(10◦C)17,6405,820840
(10◦C)0.230.06–
285384
445286
0.23 (43◦C)0.23 (60◦C)0.23 (80◦C)
D’Amico et al. 2003
CellulasePsychrophileMesophile
(4◦C)110.6
(4◦C)6.01.5
3756
––
(45◦C)40Unaffected
Garsoux et al. 2004
AminopeptidasePsychrophileMesophile
(10◦C)950114
––
3949
4758
(46◦C)1100,000
Huston et al. 2008
ImidasePsychrophileMesophile
(25◦C)25,7001,500
(25◦C)1.61.0
55>65
––
(40◦C)1502,880
Huang & Yang 2003
Lactate dehydrogenasePsychrophileThermophile
13,800 (0◦C)105,000 (44◦C)40,500 (90◦C)
0.16 (0◦C)0.41 (44◦C)0.16 (90◦C)
5090
5090
–––
Coquelle et al. 2007
Alkaline phosphatasePsychrophileMesophile
(37◦C)48,7406,954
(37◦C)0.130.11
4056
––
(50◦C)1038
Siddiqui et al. 2004b
akcat , turnover number of substrate molecules per minute per active site. Km, affinity for substrate; lower values imply higher binding affinity. Topt
(optimum temperature), temperature at which maximum enzyme activity is observed. Tm (melting temperature), temperature at which 50% of theprotein structure is in an unfolded state. t1/2 (half-life of inactivation), time needed to lose 50% of the enzyme activity at a specified temperature. Dashesindicate data not available.
(TS#). For example, reducing the activation energy from 70 kJ mol−1 for a thermophilic α-amylaseto 35 kJ mol−1 for a psychrophilic α-amylase enhanced kcat by 21-fold at 10◦C (D’Amico et al.2003). To aid substrate binding at a low energy cost, the active sites of cold-active enzymes tendto be larger and more accessible to substrates. As a result, the binding affinity of substrates forcold-active enzymes is generally lower (higher Km) than that of their thermophilic counterparts(Siddiqui & Cavicchioli 2006).
High rates of catalysis at low temperatures are generally achieved by the flexible structure andconcomitant low stability of cold-active enzymes, which is referred to as an activity-stability trade-off (Siddiqui & Cavicchioli 2006) (Table 2). Many cold-active enzymes have a more labile andflexible catalytic region than does the remainder of the protein structure, i.e., localized flexibility(Siddiqui et al. 2005, Feller 2008). Accordingly, in an environment characterized by low kineticenergy and retarded molecular motion, cold-active enzymes rely on greater disorder as a means ofmaintaining molecular dynamics and, hence, function (Feller 2007). For example, a psychrophilicalanine racemase that is very active at low temperatures and has very low thermal stability wasfound to have a hydrophilic region located at a surface loop surrounding the active site (Okubo et al.1999). The surface hydrophilic and polar regions are likely to promote solvent interactions, therebyreducing compactness and destabilizing the enzyme (Okubo et al. 1999, Siddiqui & Cavicchioli2006).
The α-amylase from P. haloplanktis, AHA, has become a model to study the structure, function,and stability relationship in cold-adapted enzymes (D’Amico et al. 2001, 2003; Feller & Gerday2003; Siddiqui & Cavicchioli 2006; Siddiqui et al. 2005, 2006; Feller 2008). Collectively, the studies
6.14 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
indicate that the structure of AHA has evolved to have relatively few electrostatic interactions inorder to provide sufficient conformational flexibility to afford activity at low temperatures, whileretaining a sufficient level of overall protein structural integrity.
Genomic analyses of psychrophilic archaea have revealed proteins characterized by a highercontent of noncharged polar amino acids (especially Gln and Thr), a lower content of hydrophobicamino acids (particularly Leu), increased exposure of hydrophobic residues, and a decreased chargethat is associated with destabilizing the surface of psychrophilic proteins (Saunders et al. 2003).Evolutionary selection of amino acid usage enabled such adaptation (Allen et al. 2009). Somewhatdifferent trends have been noted via genome surveys of marine Gammaproteobacteria where cold-adapted strains were reported to have lower contents of Ala, Arg, and Pro as well as higher contentsof Ile, Lys, and Asn (Zhao et al. 2010). Among these, Pro and Arg are associated with an abilityto confer increased stability by restricting backbone rotations and by forming multiple hydrogenbonds and salt bridges, respectively (Feller & Gerday 2003).
Psychrophilic proteins are characterized by decreased core hydrophobicity, increased surfacehydrophobicity, increased surface hydrophilicity, a lower arginine/lysine ratio, weaker interdo-main and intersubunit interactions, more and longer loops, decreased secondary structure con-tent, more glycine residues, fewer prolines in loops, more prolines in α-helices, fewer and weakermetal-binding sites, fewer disulfide bridges, fewer electrostatic interactions (H-bonds, salt bridges,cation-pi interactions, aromatic-aromatic interactions), reduced oligomerization, and an increasein the conformational entropy of the unfolded state (Siddiqui & Cavicchioli 2006). Some cold-adapted proteins also tend to have flexible 5-turn and strand secondary structures, and they possesslarge cavities lined predominantly by acidic residues to accommodate water molecules (Paredeset al. 2011). However, although the abovementioned structural features can be associated withpsychrophilic proteins, any one protein will have a limited number of, and specific context for,these structural features (Siddiqui & Cavicchioli 2006).
Other Factors Influencing Enzyme Adaptation
A cell’s cytoplasm contains high concentrations of both low- and high-molecular-weight com-pounds that lead to molecular crowding (Chebotareva et al. 2004), and under natural environ-mental conditions, microorganisms are often exposed to more than one abiotic constraint (seealso Physiological Adaptations in Psychrophiles, above). Consistent with this, the stability andactivity of enzymes are affected by the presence of organic solutes (amino acids and sugars) andpolymers (proteins and polysaccharides) (Thomas et al. 2001, Siddiqui et al. 2002, Somero 2003,Faria et al. 2008), protein-protein interactions (Thomas et al. 2001), viscosity of the intracellularand extracellular environment (Demchenko et al. 1989, Siddiqui et al. 2004a, Karan et al. 2012),and the combined effects of temperature and pressure (Saito & Nakayama 2004, Kato et al. 2008)or temperature and salt (Srimathi et al. 2007, Yan et al. 2009).
A limited number of heat-labile enzymes can also be cold-labile enzymes near or below subzerotemperatures (D’Amico et al. 2003, Xu et al. 2003), and some oligomeric or cofactor that requiresenzymes (e.g., tryptophanase) can be reversibly inactivated at lower temperatures as a result ofsubunit and cofactor dissociation (Kogan et al. 2009). Therefore, if a key cellular enzyme is coldinactivated or cold denaturated, it could define the lower temperature limit for growth rather thanthe freezing point of the aqueous environment in which the organism grows.
COLD-ADAPTED ENZYMES AND CLIMATE CHANGE
A major source of CO2 input into the atmosphere is caused by the microbial decomposition of soilorganic matter (SOM) (German et al. 2012). Predictions are that the carbon sequestered in SOM
www.annualreviews.org • Psychrophiles 6.15
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
is at least four times higher than the carbon content in the atmosphere and living plants. Globalwarming has a particularly strong effect on polar and alpine environments, wherein ∼30% of theglobal soil carbon pool resides. The degradation of cellulose, hemicellulose, and humic substancesin SOM by extracellular enzymes (e.g., glucanases, ligninases) into dissolved organic compoundsrepresents the rate-limiting step in carbon release (Weedon et al. 2011, German et al. 2012).The kinetic and thermodynamic properties of extracellular enzymes, including their responsesto environmental factors (e.g., nutrient supply, nitrogen and oxygen availability, phenolics andsubstrate concentration, soil moisture, permafrost melting, and temperature), are now beginningto be incorporated into predictive models describing the effects of global warming on carboncycling (Davidson & Janssens 2006, Weedon et al. 2011, German et al. 2012).
In view of such issues associated with global warming, it is important to recognize that cold-adapted enzymes work efficiently at low temperatures and therefore help to reduce CO2 emissionsby reducing electricity consumption associated with heating (Cavicchioli et al. 2002, 2011). Forexample, washing machines utilize a high proportion of a household’s electricity budget, and∼80% of the electricity is used to heat water (Nielsen 2005). Using cold-active enzymes, washingtemperatures can be reduced from 40◦C to 30◦C, resulting in a 30% decrease in electricity usage.Importantly, washing temperatures set 10◦C lower reduces the CO2 emissions associated with theburning of fossil fuels for energy generation by 100 g per wash (Nielsen 2005). The application ofcold-adapted enzymes in a range of other industries such as textile, food, waste-water treatment,and paper and pulp also helps to reduce toxic by-products, electricity usage, and CO2 emissions(EuropaBio Rep. 2009, Cavicchioli et al. 2011).
MICROBIAL EXTREMES, COLD-ACTIVE ENZYMES,AND ASTROBIOLOGY
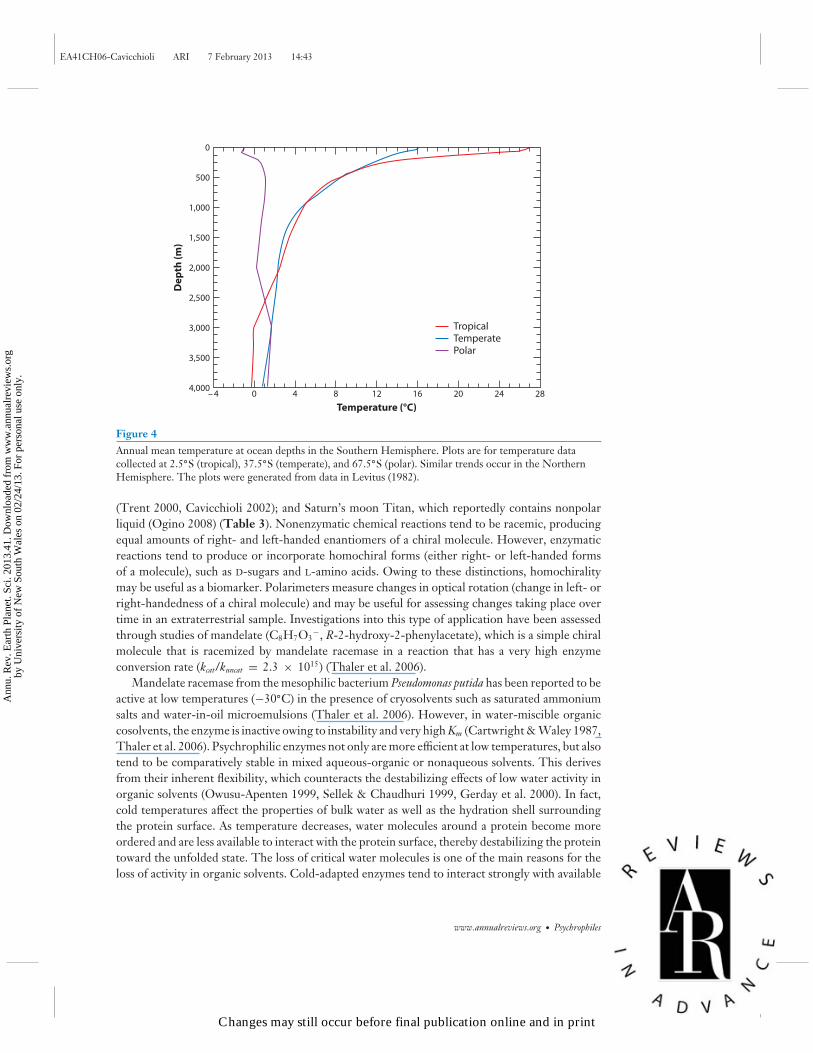
The deep sea offers a unique perspective on cold environments (Figure 4), but more manned expe-ditions to outer space have been performed than trips to the deepest reaches of the ocean. There-fore, the experience gained in overcoming issues with deep-sea exploration may translate to thedevelopment of tractable systems for biological exploration of extraterrestrial environments. Sam-pling cold deep-sea environments is logistically challenging, particularly at depths below 6,000–8,000 m, where the length of wire cable that can be carried on an oceanographic vessel is exceeded(Lauro & Bartlett 2008). As a result, in addition to the use of cable-tethered Niskin bottles for sam-ple collection (Martin-Cuadrado et al. 2007), autonomous underwater vehicles (e.g., Takami et al.1997) and free vehicles (e.g., Eloe et al. 2011b) have been developed. Arising from a limited numberof molecular studies that have been performed using such sampling designs (DeLong et al. 2006;Lauro & Bartlett 2008; Brown et al. 2009; Agogue et al. 2011; Eloe et al. 2010, 2011a), a high level ofmicrobial diversity has been identified in the deep sea. The microbiota include bacterial members ofAlpha-, Beta-, Delta-, Epsilon-, and Gammaproteobacteria as well as Actinobacteria, Bacteroidetes, Chlo-roflexi, Planctomycetes, and Verrucomicrobia. Also included are archaeal members of EuryarchaeotaMarine Groups II and III, Crenarchaeota Marine Group I, Methanopyri, and novel alveolate GroupsI and II of eucarya that include endoparasitic dinoflagellates. The capacity of microorganisms tothrive under a range of combined extremes, such as in the deep sea where adaptation to cold, highhydrostatic pressure, and nutrient limitation is required, broadens the horizons for the scope oflocations that may be considered in the search for extraterrestrial life (Cavicchioli 2002).
Cold-active enzymes may be useful for specific applications in studies aimed at searching forsigns of life in extraterrestrial environments where liquid water is known or inferred to exist,such as on Saturn’s (Enceladus) and Jupiter’s (Europa, Ganymede) icy moons; possibly Mars
6.16 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.
EA41CH06-Cavicchioli ARI 7 February 2013 14:43
0
500
1,000
1,500
2,000
2,500
3,000
3,500
4,000– 4 0 4 8
PolarTemperateTropical
12 16 20 24 28
De
pth
(m
)
Temperature (°C)
Figure 4Annual mean temperature at ocean depths in the Southern Hemisphere. Plots are for temperature datacollected at 2.5◦S (tropical), 37.5◦S (temperate), and 67.5◦S (polar). Similar trends occur in the NorthernHemisphere. The plots were generated from data in Levitus (1982).
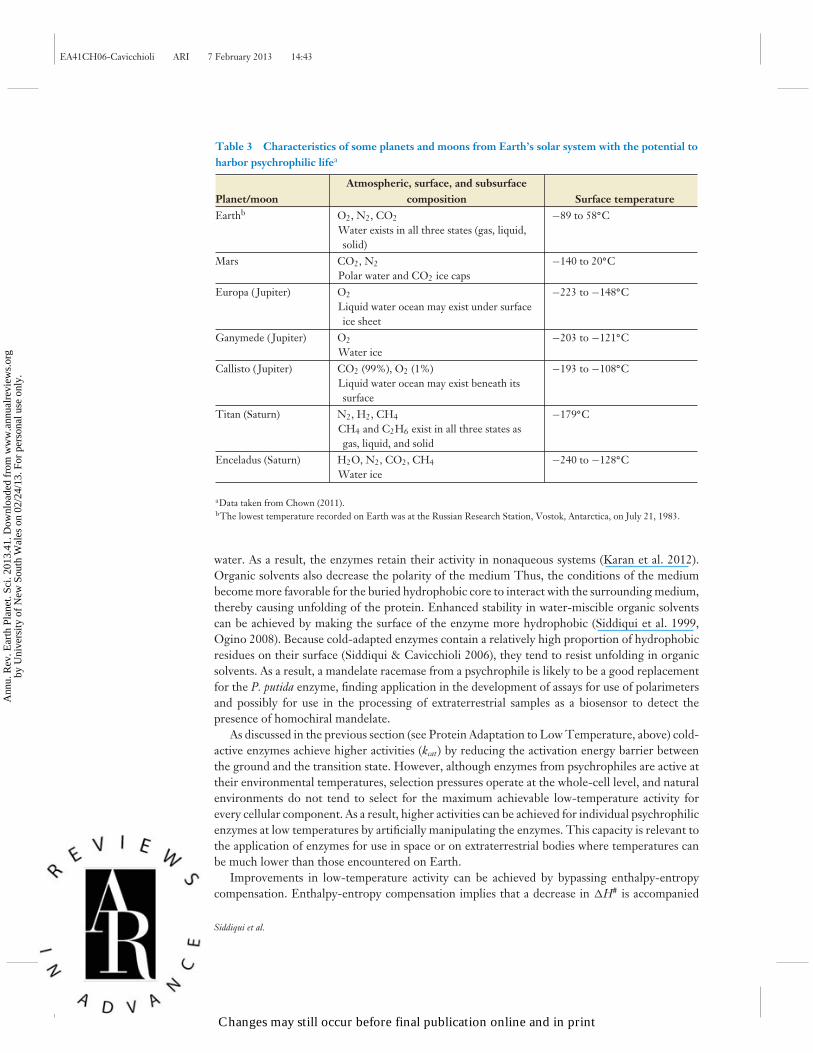
(Trent 2000, Cavicchioli 2002); and Saturn’s moon Titan, which reportedly contains nonpolarliquid (Ogino 2008) (Table 3). Nonenzymatic chemical reactions tend to be racemic, producingequal amounts of right- and left-handed enantiomers of a chiral molecule. However, enzymaticreactions tend to produce or incorporate homochiral forms (either right- or left-handed formsof a molecule), such as D-sugars and L-amino acids. Owing to these distinctions, homochiralitymay be useful as a biomarker. Polarimeters measure changes in optical rotation (change in left- orright-handedness of a chiral molecule) and may be useful for assessing changes taking place overtime in an extraterrestrial sample. Investigations into this type of application have been assessedthrough studies of mandelate (C8H7O3
−, R-2-hydroxy-2-phenylacetate), which is a simple chiralmolecule that is racemized by mandelate racemase in a reaction that has a very high enzymeconversion rate (kcat/kuncat = 2.3 × 1015) (Thaler et al. 2006).
Mandelate racemase from the mesophilic bacterium Pseudomonas putida has been reported to beactive at low temperatures (−30◦C) in the presence of cryosolvents such as saturated ammoniumsalts and water-in-oil microemulsions (Thaler et al. 2006). However, in water-miscible organiccosolvents, the enzyme is inactive owing to instability and very high Km (Cartwright & Waley 1987,Thaler et al. 2006). Psychrophilic enzymes not only are more efficient at low temperatures, but alsotend to be comparatively stable in mixed aqueous-organic or nonaqueous solvents. This derivesfrom their inherent flexibility, which counteracts the destabilizing effects of low water activity inorganic solvents (Owusu-Apenten 1999, Sellek & Chaudhuri 1999, Gerday et al. 2000). In fact,cold temperatures affect the properties of bulk water as well as the hydration shell surroundingthe protein surface. As temperature decreases, water molecules around a protein become moreordered and are less available to interact with the protein surface, thereby destabilizing the proteintoward the unfolded state. The loss of critical water molecules is one of the main reasons for theloss of activity in organic solvents. Cold-adapted enzymes tend to interact strongly with available
www.annualreviews.org • Psychrophiles 6.17
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.
EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Table 3 Characteristics of some planets and moons from Earth’s solar system with the potential toharbor psychrophilic lifea
Planet/moonAtmospheric, surface, and subsurface
composition Surface temperatureEarthb O2, N2, CO2
Water exists in all three states (gas, liquid,solid)
−89 to 58◦C
Mars CO2, N2
Polar water and CO2 ice caps−140 to 20◦C
Europa ( Jupiter) O2
Liquid water ocean may exist under surfaceice sheet
−223 to −148◦C
Ganymede ( Jupiter) O2
Water ice−203 to −121◦C
Callisto ( Jupiter) CO2 (99%), O2 (1%)Liquid water ocean may exist beneath itssurface
−193 to −108◦C
Titan (Saturn) N2, H2, CH4
CH4 and C2H6 exist in all three states asgas, liquid, and solid
−179◦C
Enceladus (Saturn) H2O, N2, CO2, CH4
Water ice−240 to −128◦C
aData taken from Chown (2011).bThe lowest temperature recorded on Earth was at the Russian Research Station, Vostok, Antarctica, on July 21, 1983.
water. As a result, the enzymes retain their activity in nonaqueous systems (Karan et al. 2012).Organic solvents also decrease the polarity of the medium Thus, the conditions of the mediumbecome more favorable for the buried hydrophobic core to interact with the surrounding medium,thereby causing unfolding of the protein. Enhanced stability in water-miscible organic solventscan be achieved by making the surface of the enzyme more hydrophobic (Siddiqui et al. 1999,Ogino 2008). Because cold-adapted enzymes contain a relatively high proportion of hydrophobicresidues on their surface (Siddiqui & Cavicchioli 2006), they tend to resist unfolding in organicsolvents. As a result, a mandelate racemase from a psychrophile is likely to be a good replacementfor the P. putida enzyme, finding application in the development of assays for use of polarimetersand possibly for use in the processing of extraterrestrial samples as a biosensor to detect thepresence of homochiral mandelate.
As discussed in the previous section (see Protein Adaptation to Low Temperature, above) cold-active enzymes achieve higher activities (kcat) by reducing the activation energy barrier betweenthe ground and the transition state. However, although enzymes from psychrophiles are active attheir environmental temperatures, selection pressures operate at the whole-cell level, and naturalenvironments do not tend to select for the maximum achievable low-temperature activity forevery cellular component. As a result, higher activities can be achieved for individual psychrophilicenzymes at low temperatures by artificially manipulating the enzymes. This capacity is relevant tothe application of enzymes for use in space or on extraterrestrial bodies where temperatures canbe much lower than those encountered on Earth.
Improvements in low-temperature activity can be achieved by bypassing enthalpy-entropycompensation. Enthalpy-entropy compensation implies that a decrease in �H# is accompanied
6.18 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
by a decrease in �S# so that an overall small increase in kcat is achieved (Siddiqui & Cavicchioli2006). However, the gain in kcat would be massive if the decrease in �H# was not accompanied bya corresponding decrease in �S# or even more so if an increase in �S# occurred. Theoretically,by maintaining a constant �S# and decreasing �H# by only 20 kJ mol−1, a 50,000-fold increasein kcat would occur at 15◦C (Lonhienne et al. 2000). Experimental work has shown that theenthalpy-entropy compensation relationship does not always hold true in cold-adapted lipasesfrom Candida antarctica, particularly in supercritical CO2 and an organic solvent (3-hexanol) wherehigher activity was associated with both negative �H# and positive �S# (Ottosson et al. 2001,2002a,b). Supercritical fluids may function as useful, nonaqueous solvents for enzyme catalysis,and they occur naturally on some planets (Mesiano et al. 1999, Comm. Origins Evol. Life Natl.Res. Counc. 2007).
kcat of a cold-adapted enzyme could be further enhanced by simultaneously decreasing �H#
and increasing �S#; this condition could be achieved on an extraterrestrial body where a water-likepolar solvent is present by indirectly increasing the entropy of the system via solvent displacement(Wolfenden & Snider 2001, Snider et al. 2002). If more solvent molecules are released uponbinding to the transition state of the enzyme than upon binding to the ground-state substrate,then there will be considerable entropic benefit for the formation of an enzyme-transition-statecomplex that has a concomitant increase in activity (Wolfenden & Snider 2001).
To design highly active enzymes from antibodies (catalytic antibodies), reaction rates canbe enhanced by promoting the release of water from the binding pocket during formation ofthe transition state and thereby producing an increase in �S# (Houk et al. 2003). Similarly,an enhanced rate of reaction for ribosome-mediated peptide bond formation can be achievedby effective substrate positioning and/or by water exclusion from within the active site, whichcreates an increase in �S# (Wolfenden 2011). Therefore, in theory, enzyme reactions, biologicalprocesses, and metabolically active life may be achievable under very cold planetary conditions,provided that a decrease in �H# is accompanied by either no change or an increase in �S# duringenzyme catalysis (i.e., surmounting enthalpy-entropy compensation).
Although this review focuses on unicellular microorganisms, as a parting note we highlightthe remarkable properties of the small (∼0.1–1 mm in length) metazoans (panarthropods) calledtardigrades (“waterbears”). Tardigrades are adapted to multiple extremes, and in both their hy-drated (active) and dehydrated (tun) forms, they are resistant to very cold temperatures. Antarctictardigrades have survived exposure to −22◦C for 600 and 3,040 days in active and tun states,respectively, with some in their tun state surviving up to 14 days at −180◦C (Somme & Meier1995). Given their tolerance to cold and other extremes, tardigrades are recognized as valuablemetazoan models for astrobiological research (Horikawa et al. 2008): They were used aboard theFOTON-M3 mission to examine their resistance to the effects of outer space (LIFE-TARSEproject) (Rebecchi et al. 2009).
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
This work was supported by the Australian Research Council and the Australian Antarctic ScienceProgram. We thank Mark Milnes for the panoramic image of Deep Lake in Figure 2.
www.annualreviews.org • Psychrophiles 6.19
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
LITERATURE CITED
Adams MWW, Kelly RM. 1994. Thermostability and thermoactivity of enzymes from hyperthermophilicarchaea. Bioorg. Med. Chem. 2:659–67
Agogue H, Lamy D, Neal PR, Sogin ML, Herndl GJ. 2011. Water mass-specificity of bacterial communitiesin the North Atlantic revealed by massively parallel sequencing. Mol. Ecol. 20:258–74
Alekhina IA, Marie D, Petit JR, Lukin VV, Zubkov VM, Bulat SA. 2007. Molecular analysis of bacterialdiversity in kerosene-based drilling fluid from the deep ice borehole at Vostok, East Antarctica. FEMSMicrobiol. Ecol. 59:289–99
Allen M, Lauro FM, Williams TJ, Burg D, Siddiqui KS, et al. 2009. The genome sequence of the psychrophilicarchaeon, Methanococcoides burtonii: the role of genome evolution in cold adaptation. ISME J. 3:1012–35
Amann R, Ludwig W, Schleifer K. 1995. Phylogenetic identification and in situ detection of individualmicrobial cells without cultivation. Microbiol. Rev. 59:143–69
Amato P, Christner BC. 2009. Energy metabolism response to low-temperature and frozen conditions inPsychrobacter cryohalolentis. Appl. Environ. Microbiol. 75:711–18
Anderson DT, Sumner DY, Hawes I, Webster-Brown J, McKay CP. 2011. Discovery of large conicalstromatolites in Lake Untersee, Antarctica. Geobiology 9:280–93
Anesio AM, Bellas CM. 2011. Are low temperature habitats hot spots of microbial evolution driven byviruses? Trends Microbiol. 19:52–57
Antibus DE, Leff LG, Hall BL, Baeseman JL, Blackwood CB. 2012a. Cultivable bacteria from ancient algalmats from the McMurdo Dry Valleys, Antarctica. Extremophiles 16:105–14
Antibus DE, Leff LG, Hall BL, Baeseman JL, Blackwood CB. 2012b. Molecular characterization of ancientalgal mats from McMurdo Dry Valleys, Antarctica. Antarctic Sci. 24:139–46
Bakermans C, Nealson KH. 2004. Relationship of critical temperature to macromolecular synthesis andgrowth yield in Psychrobacter cryopegella. J. Bacteriol. 186:2340–45
Bakermans C, Tollaksen SL, Giometti CS, Wilkerson C, Tiedje JM, et al. 2007. Proteomic analysis ofPsychrobacter cryohalolentis K5 during growth at subzero temperatures. Extremophiles 11:343–54
Bergholz PW, Bakermans C, Tiedje JM. 2009. Psychrobacter arcticus 273–4 uses resource efficiency andmolecular motion adaptations for subzero temperature growth. J. Bacteriol. 191:2340–52
Bielewicz S, Bell E, Kong W, Friedberg I, Priscu JC, Morgan-Kiss RM. 2011. Protist diversity in apermanently ice-covered Antarctic Lake during the polar night transition. ISME J. 5:1559–64
Bomblies A, McKnight DM, Andrews ED. 2001. Retrospective simulation of lake-level rise in Lake Bonneybased on recent 21-year record: indication of recent climate change in the McMurdo Dry Valleys,Antarctica. J. Paleolimnol. 25:477–92
Borghini F, Colacevich A, Bargagli R. 2010. A study of autotrophic communities in two Victoria Land lakes(Continental Antarctica) using photosynthetic pigments. J. Limnol. 69:333–40
Bowman JP, McCammon SA, Brown JL, McMeekin TA. 1998a. Glaciecola punicea gen. nov., sp. nov. andGlaciecola pallidula gen. nov., sp. nov.: psychrophilic bacteria from Antarctic sea-ice habitats. Int. J. Syst.Bacteriol. 48:1213–22
Bowman JP, McCammon SA, Brown JL, Nichols PD, McMeekin TA. 1997a. Psychroserpens burtonensis gen.nov., sp. nov., and Gelidibacter algens gen. nov., sp. nov., psychrophilic bacteria isolated from Antarcticlacustrine and sea ice habitats. Int. J. Syst. Bacteriol. 14:670–77
Bowman JP, McCammon SA, Brown MV, Nichols DS, McMeekin TA. 1997b. Diversity and association ofpsychrophilic bacteria in Antarctic sea ice. Appl. Environ. Microbiol. 63:3068–78
Bowman JP, McCammon SA, Gibson JA, Robertson L, Nichols PD. 2003. Prokaryotic metabolic activityand community structure in Antarctic continental shelf sediments. Appl. Environ. Microbiol. 69:2448–62
Bowman JP, McCammon SA, Lewis T, Skerratt JH, Brown JL, et al. 1998b. Psychroflexus torquis gen. nov.,sp. nov., a psychrophilic species from Antarctic sea ice and reclassification of Flavobacterium gondwanense(Dobson et al. 1993) as Psychroflexus gondwanense gen. nov., comb. nov. Microbiology 144:1601–9
Bowman JP, McCammon SA, Rea SM, McMeekin TA. 2000a. The microbial composition of threelimnologically disparate hypersaline Antarctic lakes. FEMS Microbiol. Lett. 183:81–88
Bowman JP, Nichols DS, McMeekin TA. 1997c. Psychrobacter glacincola sp. nov., a halotolerant, psychrophilicbacterium isolated from Antarctic sea ice. Syst. Appl. Microbiol. 20:209–15
6.20 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Bowman JP, Rea SM, McCammon SA, McMeekin TA. 2000b. Diversity and community structure withinanoxic sediment from marine salinity meromictic lakes and a coastal meromictic marine basin, VestfoldHills, Eastern Antarctica. Environ. Microbiol. 2:227–37
Brambilla E, Hippe H, Hagelstein A, Tindall BJ, Stackebrandt E. 2001. 16S rDNA diversity of culturedand uncultured prokaryotes of a mat samples from Lake Fryxell, McMurdo Dry Valleys, Antarctica.Extremophiles 5:23–33
Brown MV, Lauro FM, DeMeare MZ, Muir L, Wilkins D, et al. 2012. Global biogeography of SAR11marine bacteria. Mol. Syst. Biol. 8:595
Brown MV, Philip GK, Bunge JA, Smith MC, Bissett A, et al. 2009. Microbial community structure in theNorth Pacific Ocean. ISME J. 3:1374–86
Callejas C, Gill PR, Catalan AI, Azziz G, Castro-Sowinski S, Batista S. 2011. Phylotype diversity in abenthic cyanobacterial mat community on King George Island, maritime Antarctica. World J. Microbiol.Biotechnol. 27:1507–12
Campbell PJ. 1978. Primary productivity of a hypersaline Antarctic lake. Aust. J. Mar. Freshw. Res. 29:717–24Cartwright SJ, Waley SG. 1987. Cryobiology of β-lactamases. Biochemistry 26:5329–37Cary CS, McDonald IR, Barrett JE, Cowan DA. 2010. On the rocks: the microbiology of Antarctic Dry
Valley soils. Nat. Rev. Microbiol. 8:129–38Casanueva A, Tuffin M, Cary C, Cowan DA. 2010. Molecular adaptations to psychrophily: the impact of
‘omic’ technologies. Trends Microbiol. 18:374–81Cavalieri DJ, Parkinson CL. 2008. Antarctic sea ice variability and trends 1979–2006. J. Geophys. Res.
113:C07004Cavicchioli R. 2002. Extremophiles and the search for extra-terrestrial life. Astrobiology 2:281–92Cavicchioli R. 2006. Cold-adapted Archaea. Nat. Rev. Microbiol. 4:331–43Cavicchioli R. 2007. Antarctic metagenomics. Microbiol. Aust. 28:98–103Cavicchioli R, Charlton T, Ertan H, Mohd Omar S, Siddiqui KS, Williams TJ. 2011. Biotechnological uses
of enzymes from psychrophiles. Microb. Biotechnol. 4:449–60Cavicchioli R, Curmi PMG, Siddiqui KS, Thomas T. 2006. Proteins from psychrophiles. In Methods in
Microbiology, Vol. 35: Extremophiles, ed. FA Rainey, A Oren, pp. 395–436. London: AcademicCavicchioli R, Siddiqui KS. 2006. Cold-adapted enzymes. In Enzyme Technology, ed. A Pandey, C Webb, CR
Soccol, C Larroche, pp. 615–38. New York: Springer Sci.Cavicchioli R, Siddiqui KS, Sowers KR, Andrews D. 2002. Low-temperature extremophiles and their
applications. Curr. Opin. Biotechnol. 13:253–61Chebotareva NA, Kurganov BI, Livanova NB. 2004. Biochemical effects of molecular crowding. Biochemistry
69:1239–51Chown M. 2011. Solar System: A Visual Exploration of the Planets, Moons, and Other Heavenly Bodies that Orbit
Our Sun. London: Faber & Faber. 224 pp.Christiansen JK, Larsen MH, Ingmer H, Søgaard-Andersen L, Kallipolitis BH. 2004. The RNA-binding
protein Hfq of Listeria monocytogenes: role in stress tolerance and virulence. J. Bacteriol. 186:3355–62Christner BC, Mosley-Thompson E, Thompson LG, Reeve JN. 2001. Isolation of bacteria and 16S rDNAs
from Lake Vostok accretion ice. Environ. Microbiol. 3:570–77Comm. Origins Evol. Life Natl. Res. Counc. 2007. The Limits of Organic Life in Planetary Systems. Washington,
DC: Natl. Acad. Press. 116 pp.Coquelle N, Fioravanti E, Weik M, Vellieux F, Madern D. 2007. Activity, stability and structural studies of
lactate dehydrogenases adapted to extreme thermal environments. J. Mol. Biol. 374:547–62Crawford DL, Powers DA. 1992. Evolutionary adaptation to different thermal environments via transcrip-
tional regulation. Mol. Biol. Evol. 9:806–13Curran MAJ, van Ommen TD, Morgan VI, Phillips KL, Palmer AS. 2003. Ice core evidence for Antarctic
sea ice decline since the 1950s. Science 302:1203–6Dalluge JJ, Hamamoto T, Horikoshi K, Morita RY, Stetter KO, McCloskey JA. 1997. Posttranscriptional
modification of tRNA in psychrophilic bacteria. J. Bacteriol. 179:1918–23D’Amico S, Collins S, Marx J-C, Feller G, Gerday C. 2006. Psychrophilic microorganisms: challenges for
life. EMBO Rep. 7:385–89
www.annualreviews.org • Psychrophiles 6.21
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
D’Amico S, Gerday C, Feller G. 2001. Structural determinants of cold adaptation and stability in a largeprotein. J. Biol. Chem. 276:25791–96
D’Amico S, Marx JC, Gerday C, Feller G. 2003. Activity-stability relationships in extremophilic enzymes. J.Biol. Chem. 278:7891–96
Daniel RM, Danson MJ, Hough DW, Lee CK, Peterson ME, et al. 2008. Enzyme stability and activity athigh temperatures. See Siddiqui & Thomas 2008, pp. 1–34
Davidson EA, Janssens IA. 2006. Temperature sensitivity of soil carbon decomposition and feedback toclimate change. Nature 440:165–73
DeLong EF, Preston CM, Mincer T, Rich V, Hallam SJ, et al. 2006. Community genomics among stratifiedmicrobial assemblages in the ocean’s interior. Science 311:496–503
Demchenko AP, Rusyn OI, Saburova EA. 1989. Kinetics of the lactate dehydrogenase reaction inhigh-viscosity media. Biochim. Biophys. Acta 998:196–203
Demirjian DC, Morıs-Varas F, Cassidy CS. 2001. Enzymes from extremophiles. Curr. Opin. Chem. Biol.5:144–51
Ehira S, Hamano T, Hayashida T, Kojima K, Nakamoto H, et al. 2003. Conserved temperature-dependentexpression of RNA-binding proteins in cyanobacteria with different temperature optima. FEMSMicrobiol. Lett. 225:137–42
Eicken H. 2003. From the microscopic, to the macroscopic, to the regional scale: growth, microstructure,and properties of sea ice. In Sea Ice: An Introduction to Its Physics, Chemistry, Biology and Geology, ed. DNThomas, GS Dieckmann, pp. 22–81. Oxford, UK: Blackwell Sci.
Elenda C, Schmeissera C, Hoebenreich H, Steelea HL, Streit WR. 2007. Isolation and characterizationof a metagenome-derived and cold-active lipase with high stereospecificity for (R)-ibuprofen esters. J.Biotechnol. 130:370–77
Eloe EA, Fadrosh DW, Novotny M, Zeigler Allen L, Kim M, et al. 2011a. Going deeper: metagenome of ahadopelagic microbial community. PLoS ONE 6:e20388
Eloe EA, Malfatti F, Gutierrez J, Hardy K, Schmidt WE, et al. 2011b. Isolation and characterization of apsychropiezophilic alphaproteobacterium. Appl. Environ. Microbiol. 77:8145–53
Eloe EA, Shulse CN, Fadrosh DW, Williamson SJ, Allen EA, et al. 2010. Compositional differences inparticle-associated and free-living microbial assemblages from an extreme deep-ocean environment.Environ. Microbiol. Rep. 3:449–58
EuropaBio Rep. 2009. How industrial biology can tackle climate change. EuropaBio, Jan. 28, Bruss., Belg. http://www.europabio.org/industrial/positions/how-industrial-biotechnology-can-tackle-climate-change
Falkowski P. 2012. The power of plankton. Nature 483:S17–20Faria TQ, Mingote A, Siopa F, Ventura R, Maycock C, Santos H. 2008. Design of new enzyme stabilizers
inspired by glycosides of hyperthermophilic microorganisms. Carbohydr. Res. 343:3025–33Feller G. 2007. Life at low temperatures: Is disorder the driving force? Extremophiles 11:211–16Feller G. 2008. Enzyme function at low temperatures in psychrophiles. See Siddiqui & Thomas 2008, pp. 35–69Feller G, Gerday C. 2003. Psychrophilic enzymes: hot topics in cold adaptation. Nat. Rev. Microbiol. 1:200–8Fernandez-Carazo R, Hodgson DA, Convey P, Wilmotte A. 2011. Low cyanobacterial diversity in biotypes
of the Transantarctic Mountains and Shackleton Range (80–82◦S), Antarctica. FEMS Microbiol. Ecol.77:503–17
Fernandez-Valiente E, Camacho A, Rochera C, Rico E, Vincent WF, Quesada A. 2007. Communitystructure and physiological characterization of microbial mats in Byers Peninsula, Livingston Island(South Shetland Islands, Antarctica). FEMS Microbiol. Ecol. 59:377–85
Ferris JM, Burton HR. 1988. The annual cycle of heat content and mechanical stability of hypersaline DeepLake, Vestfold Hills, Antarctica. Hydrobiologia 165:115–28
Franzmann PD, Liu Y, Balkwill DL, Aldrich HC, Conway de Macario E, Boone DR. 1997. Methanogeniumfrigidum sp. nov., a psychrophilic, H2-using methanogen from Ace Lake, Antarctica. Int. J. Syst. Bacteriol.47:1068–72
Franzmann PD, Stackebrandt E, Sanderson K, Volkman JK, Cameron DE, et al. 1988. Halobacterium lacuspro-fundi sp. nov., a halophilic bacterium isolated from Deep Lake, Antarctica. Syst. Appl. Microbiol. 11:20–27
6.22 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Franzmann PD, Stringer N, Ludwig W, Conway de Macario E, Rohde M. 1992. A methanogenic archaeonfrom Ace Lake, Antarctica: Methanococcoides burtonii sp. nov. Syst. Appl. Microbiol. 15:573–81
Gao H, Yanh ZK, Wu L, Thompson DK, Zhou J. 2006. Global transcriptome analysis of the cold shockresponse of Shewanella oneidensis MR-1 and mutational analysis of its classical cold shock proteins.J. Bacteriol. 188:4560–69
Garsoux G, Lamotte J, Gerday C, Feller G. 2004. Kinetic and structural optimization to catalysis at lowtemperatures in a psychrophilic cellulase from the Antarctic bacterium Pseudoalteromonas haloplanktis.Biochem. J. 384:247–53
Gerday C, Aittaleb M, Bentahir M, Chessa JP, Claverie P, et al. 2000. Cold-adapted enzymes: fromfundamentals to biotechnology. Trends Biotechnol. 18:103–7
German DP, Marcelo KRB, Stone MM, Allison SD. 2012. The Michaelis-Menten kinetics of soil extracellularenzymes in response to temperature: a cross-latitudinal study. Glob. Change Biol. 18:1468–79
Giaquinto L, Curmi PM, Siddiqui KS, Poljak A, DeLong E, et al. 2007. Structure and function of cold shockproteins in archaea. J. Bacteriol. 189:5738–48
Gibson JAE. 1999. The meromictic lakes and stratified marine basins of the Vestfold Hills, East Antarctica.Antarctic Sci. 11:175–92
Gibson JAE. 2006. Limnology of epiglacial lakes of the Framnes Mountains, Antarctica: insights into a widespread butpoorly studied lake type. Presented at Sci. Comm. Antarctic Res. (SCAR) Open Sci. Conf., XXIX, Hobart,Aust.
Gibson JAE, Miller MR, Davies NW, Neill GP, Nichols DS, Volkman JK. 2005. Unsaturated diether lipidsin the psychrotrophic archaeon Halorubrum lacusprofundi.Syst. Appl. Microbiol. 28:19–26
Glatz RE, Lepp PW, Ward BB, Francis CA. 2006. Planktonic microbial community composition across steepphysical/chemical gradients in permanently ice-covered Lake Bonney, Antarctica. Geobiology 4:53–67
Goodchild A, Saunders NFW, Ertan H, Raftery M, Guilhaus M, et al. 2004. A proteomic determination ofcold adaptation in the Antarctic archaeon, Methanococcoides burtonii. Mol. Microbiol. 53:309–21
Gordon DA, Priscu J, Giovannoni S. 2000. Origin and phylogeny of microbes living in permanent Antarcticlake ice. Microb. Ecol. 39:197–202
Gramling C. 2012. A tiny window opens into Lake Vostok, while a vast continent awaits. Science 335:788–89Grzymski JJ, Riesenfeld CS, Williams TJ, Dussaq AM, Ducklow H, et al. 2012. A metagenomic assessment of
winter and summer bacterioplankton from Antarctic Peninsula coastal surface waters. ISME J. 6:1901–15Hawes I, Sumner DY, Anderson DT, Mackey TJ. 2011. Legacies of recent environmental change in the
benthis communities of Lake Joyce, a perennially ice-covered Antarctic lake. Geobiology 9:394–410Hebraud M, Potier P. 1999. Cold shock response and low temperature adaptation in psychrotrophic bacteria.
J. Mol. Microbiol. Biotechnol. 1:211–19Hogg OT, Barnes DKA, Griffiths HJ. 2011. Highly diverse, poorly studied and uniquely threatened by climate
change: an assessment of marine biodiversity on South Georgia’s continental shelf. PLoS ONE 6:e19795Horikawa DD, Kunieda T, Abe W, Watanabe M, Nakahara Y, et al. 2008. Establishment of a rearing
system of the extremotolerant tardigrade Ramazzottius varieornatus: a new model animal for astrobiology.Astrobiology 8:549–56
Houk KN, Leach AG, Kim SP, Zhang X. 2003. Binding affinities of host-guest, protein-ligand, andprotein-transition-state complexes. Angew. Chem. Int. Ed. Engl.42:4872–97
Huang CY, Yang YS. 2003. A novel cold-adapted imidase from fish Oreochromis niloticus that catalyzeshydrolysis of maleimide. Biochem. Biophys. Res. Commun. 312:467–72
Huston AL, Haeggstrom JZ, Feller G. 2008. Cold adaptation of enzymes: structural, kinetic and microcalori-metric characterizations of an aminopeptidase from the Arctic psychrophile Colwellia psychrerythraea andof human leukotriene A4 hydrolase. Biochim. Biophys. Acta 1784:1865–72
Inman M. 2005. Antarctic drilling: the plan to unlock Lake Vostok. Science 310:611–12Jones N. 2012. Russians celebrate Vostok victory. Nature 482:287Jones PG, Inouye M. 1994. The cold-shock response: a hot topic. Mol. Microbiol. 11:811–18Jung YH, Yi JY, Jung HJ, Lee YK, Lee HK, et al. 2010. Overexpression of cold shock protein A of Psychromonas
arctica KOPRI 22215 confers cold resistance. Protein J. 29:136–42
www.annualreviews.org • Psychrophiles 6.23
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Jungblut A-D, Hawes I, Mountfort D, Hitzfeld B, Dietrich DR, et al. 2005. Diversity within cyanobacterialmat communities in variable salinity meltwater ponds of McMurdo Ice Shelf, Antarctica. Environ.Microbiol. 7:519–29
Junge K, Eicken H, Deming JW. 2004. Bacterial activity at −2 to −20◦C in Arctic wintertime sea ice. Appl.Environ. Microbiol. 70:550–57
Karan R, Capes MD, DasSarma S. 2012. Function and biotechnology of extremophilic enzymes in low wateractivity. Aquat. Biosyst. 8:4
Karr EA, Ng JM, Belchik SM, Sattley WM, Madigan MT, Achenbach LA. 2006. Biodiversity of methanogenicand other Archaea in the permanently frozen Lake Fryxell. Appl. Environ. Microbiol. 72:1663–66
Karr EA, Sattley WM, Jung DO, Madigan MT, Achenbach LA. 2003. Remarkable diversity of phototrophicpurple bacteria in a permanently frozen Antarctic lake. Appl. Environ. Microbiol. 69:4910–14
Karr EA, Sattley WM, Rice MR, Jung DO, Madigan MT, et al. 2005. Diversity and distribution ofsulfate-reducing bacteria in permanently frozen Lake Fryxell, McMurdo Dry Valleys, Antarctica. Appl.Environ. Microbiol. 71:6353–59
Kato C, Sato T, Abe F, Ohmae E, Tamegai H, et al. 2008. Protein adaptation to high-pressure environments.See Siddiqui & Thomas 2008, pp. 167–91
Kepner RL, Wharton RA, Suttle CA. 1998. Viruses in Antarctic lakes. Limnol. Oceanogr. 43:1754–61Kim EY, Oh KH, Lee MH, Kang CH, Oh TK, et al. 2009. Novel cold-adapted alkaline lipase from an intertidal
flat metagenome and proposal for a new family of bacterial lipases. Appl. Environ. Microbiol. 75:257–60Kintisch E, Stoksta E. 2008. Ocean CO2 studies look beyond coral. Science 319:1029Kirchman DL, Moran XAG, Ducklow H. 2009. Microbial growth in the polar oceans: role of temperature
and potential impact of climate change. Nat. Rev. Microbiol. 7:451–59Kogan A, Gdalevsky GY, Cohen-Luria R, Goldgur Y, Phillips RS, et al. 2009. Conformational changes and
loose packing promote E. coli Tryptophanase cold lability. BMC Struct. Biol. 9:65Kurosawa N, Sato S, Kawarabayasi Y, Imura S, Naganuma T. 2010. Archaeal and bacterial community
structures in the anoxic sediment of Antarctic meromictic lake Nurume-Ike. Polar Sci. 4:421–29Lauro FM, Allen M, Wilkins D, Williams TJ, Cavicchioli R. 2011a. Genetics, genomics and evolution of
psychrophiles. In Extremophiles Handbook, ed. K Horikoshi, G Antranikian, F Robb, K Stetter, AT Bull,pp. 1179–99. Heidelberg: Springer
Lauro FM, Bartlett DH. 2008. Prokaryotic lifestyles in deep sea habitats. Extremophiles 12:15–25Lauro FM, DeMaere MZ, Yau S, Brown MV, Ng C, et al. 2011b. An integrative study of a meromictic lake
ecosystem in Antarctica. ISME J. 5:879–95Laybourn-Parry J, Hofer JS, Sommaruga R. 2001. Viruses in the plankton of freshwater and saline Antarctic
lakes. Freshw. Biol. 46:1279–87Laybourn-Parry J, Marshall WA, Madan NJ. 2007. Viral dynamics and patterns of lysogeny in saline Antarctic
lakes. Polar Biol. 30:351–58Laybourn-Parry J, Pearce D. 2007. The biodiversity and ecology of Antarctic lakes: models for evolution.
Philos. Trans. R. Soc. Lond. B 364:2273–89Le Quere C, Rodenbeck C, Buitenhuis ET, Conway TJ, Langenfelds R, et al. 2007. Saturation of the
Southern Ocean CO2 sink due to recent climate change. Science 136:1735–38Levitus S. 1982. Climatological Atlas of the World Oceans. NOAA Prof. Pap. No. 13. Washington, DC: U.S.
GPO. 173 pp.Lim J, Thomas T, Cavicchioli R. 2000. Low temperature regulated DEAD-box RNA helicase from the
Antarctic archaeon, Methanococcoides burtonii. J. Mol. Biol. 297:553–67Liu J, Curry JA. 2010. Accelerated warming of the Southern Ocean and its impacts on the hydrological cycle
and sea ice. Proc. Natl. Acad. Sci. USA 107:14987–92Lonhienne T, Gerday C, Feller G. 2000. Psychrophilic enzymes: revisiting the thermodynamic parameters
of activation may explain local flexibility. Biochim. Biophys. Acta 1543:1–10Lopez-Bueno A, Tamames J, Velazquez D, Moya A, Quesada A, Alcami A. 2009. High diversity of the viral
community from an Antarctic lake. Science 326:858–61Lukin V, Bulat S. 2011. Vostok subglacial lake: details of Russian plans/activities for drilling and sampling.
In Antarctic Subglacial Aquatic Environments, Geophys. Monogr. Ser., ed. MJ Siegert, MC Kennicutt II,RA Bindschadler, 192:187–97. Washington, DC: AGU
6.24 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Madan NJ, Marshall WA, Laybourn-Parry J. 2005. Virus and microbial loop dynamics over an annual cyclein three contrasting Antarctic lakes. Freshw. Biol. 50:1291–300
Mancuso Nichols C, Lardiere SG, Bowman JP, Nichols PD, Gibson JAE, Guezennec J. 2005a. Chemicalcharacterization of exopolysaccharides from Antarctic marine bacteria. Microb. Ecol. 49:578–89
Mancuso Nichols CA, Guezennec J, Bowman JP. 2005b. Bacterial exopolysaccharides from extreme marineenvironments with special consideration of the Southern Ocean, sea ice, and deep-sea hydrothermalvents: a review. Mar. Biotechnol. 7:253–71
Margesin R, Miteva V. 2011. Diversity and ecology of psychrophilic microorganisms. Res. Microbiol.162:346–61
Martın-Cuadrado AB, Lopez-Garcıa P, Alba JC, Moreira D, Monticelli L, et al. 2007. Metagenomics of thedeep Mediterranean, a warm bathypelagic habitat. PLoS ONE 2:e914
Maruyama K, Sato N, Ohta N. 1999. Conservation of structure and cold-regulation of RNA-binding proteinsin Cyanobacteria: probable convergent evolution with eukaryotic glycine-rich RNA-binding proteins.Nucleic Acids Res. 27:2029–36
Matear RJ, Hirst AC. 1999. Climate change feedback on the future oceanic CO2 uptake. Tellus 51:722–33Matsuzaki M, Kubota K, Satoh T, Kunugi M, Ban S, et al. 2006. Dimethyl sulfoxide-respiring bacteria in
Suribati Ike, a hypersaline lake, in Antarctic and the marine environment. Polar Biosci. 20:73–81McNeil BI, Matear RJ. 2008. Southern Ocean acidification: a tipping point at 450-ppm atmospheric CO2.
Proc. Natl. Acad. Sci. USA 105:18860–64Meredith MP, King JC. 2005. Rapid climate change in the ocean west of the Antarctic Peninsula during the
second half of the 20th century. Geophys. Res. Lett. 32:L19604Mesiano AJ, Beckman EJ, Russell AJ. 1999. Supercritical biocatalysis. Chem. Rev. 99:623–34Mikaloff Fletcher SE, Gruber N, Jacobson AR, Doney SC, Dutkiewicz S, et al. 2006. Inverse estimates of
anthropogenic CO2 uptake, transport, and storage by the ocean. Glob. Biogeochem. Cycles 20:1–16Mikucki JA, Pearson A, Johnston DT, Turchyn AV, Farquhar J, et al. 2009. A contemporary microbially
maintained subglacial ferrous “ocean.” Science 324:397–400Mikucki JA, Priscu JC. 2007. Bacterial diversity associated with Blood Falls, a subglacial outfile from the
Taylor Glacier, Antarctic. Appl. Environ. Microbiol. 73:4029–39Mock T, Thomas DN. 2005. Recent advances in sea-ice microbiology. Environ. Microbiol. 7:605–19Moline MA, Claustre H, Frazer TK, Schofield O, Vernet M. 2004. Alteration of the food web along the
Antarctic Peninsula in response to a regional warming trend. Glob. Change Biol. 10:1973–80Moorhead D, Schmeling J, Hawes I. 2005. Modelling the contribution of benthic microbial mats to net
primary production in Lake Hoare, McMurdo Dry Valleys. Antarctic Sci. 17:33–45Mori S, Castoreno A, Mulligan ME, Lammers PJ. 2003. Nitrogen status modulates the expression of
RNA-binding proteins in cyanobacteria. FEMS Microbiol. Lett. 227:203–10Mosier AC, Murray AE, Fritsen CH. 2007. Microbiota within the perennial ice cover of Lake Vida, Antarctica.
FEMS Microbiol. Ecol. 59:274–88Murray AE, Grzymski JJ. 2007. Diversity and genomics of Antarctic marine micro-organisms. Philos. Trans.
R. Soc. Lond. B 362:2259–71Ng C, DeMaere MZ, Williams TJ, Lauro FM, Raftery M, et al. 2010. Metaproteogenomic analysis of a
dominant green sulfur bacterium from Ace Lake, Antarctica. ISME J. 4:1002–19Nichols DS, Miller MR, Davies NW, Goodchild A, Raftery M, Cavicchioli R. 2004. Cold adaptation in
the Antarctic Archaeon Methanococcoides burtonii involves membrane lipid unsaturation. J. Bacteriol.186:8508–15
Nielsen PH. 2005. Life cycle assessment supports cold-wash enzymes. SOFW-J. 10:1–4Noon KR, Guymon R, Crain PF, McCloskey JA, Thomm M, et al. 2003. Influence of temperature on tRNA
modification in Archaea: Methanococcoides burtonii (optimum growth temperature [Topt], 23◦C) andStetteria hydrogenophila (Topt, 95◦C). J. Bacteriol. 185:5483–90
Ogino H. 2008. Organic solvent-stable enzymes. See Siddiqui & Thomas 2008, pp. 193–36Okubo Y, Yokoigawa K, Esaki N, Soda K, Kawai H. 1999. Characterization of psychrophilic alanine racemase
from Bacillus psychrosaccharolyticus. Biochem. Biophys. Res. Commun. 256:333–40Olsen JB, Steppe TF, Litaker RW, Paerl HW. 1998. N2-fixing microbial consortia associated with the ice
cover of Lake Bonney, Antarctica. Microb. Ecol. 36:231–38
www.annualreviews.org • Psychrophiles 6.25
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Ottosson J, Fransson L, Hult K. 2002a. Substrate entropy in enzyme enantioselectivity: an experimental andmolecular modeling study of a lipase. Protein Sci. 11:1462–71
Ottosson J, Fransson L, King JW, Hult K. 2002b. Size as a parameter for solvent effects on Candida antarcticalipase B enantioselectivity. Biochim. Biophys. Acta 1594:325–34
Ottosson J, Rotticci-Mulder JC, Rotticci D, Hult K. 2001. Rational design of enantioselective enzymesrequires considerations of entropy. Protein Sci. 10:1769–74
Owusu-Apenten RK. 1999. Low temperature organic phase biocatalysis using cold-adapted enzymes. InBiotechnological Applications of Cold-Adapted Organisms, ed. R Margesin, F Schinner, pp. 35–48. Heidelberg:Springer
Paredes DI, Watters K, Pitman DJ, Bystroff C, Dordick JS. 2011. Comparative void-volume analysis ofpsychrophilic and mesophilic enzymes: structural bioinformatics of psychrophilic enzymes revealssources of core flexibility. BMC Struct. Biol. 11:42
Pearce DA. 2003. Bacterioplankton community structure in a maritime Antarctic oligotrophic lake during aperiod of holomixis, as determined by denaturing gradient gel electrophoresis (DGGE) and fluorescencein situ hybridization (FISH). Microb. Ecol. 46:92–105
Pearce DA. 2005. The structure and stability of the bacterioplankton community in Antarctic freshwaterlakes, subject to extremely rapid environmental change. FEMS Microb. Ecol. 53:61–72
Pearce DA, van der Gast CJ, Lawley B, Ellis-Evans JC. 2003. Bacterioplankton community diversity in amaritime Antarctic lake, determined by culture-dependent and culture-independent techniques. FEMSMicrobiol. Ecol. 45:59–70
Pearce DA, van der Gast CJ, Woodward K, Newsham KK. 2005. Significant changes in the bacterioplanktoncommunity structure of a maritime Antarctic freshwater lake following nutrient enrichment. Microbiology151:3237–48
Peeters K, Hodgson DA, Convey P, Willems A. 2011. Culturable diversity of heterotrophic bacteria inForlidas Pond (Pensacola Mountains) and Lundstrom Lake (Shackleton Range), Antarctica. Microb. Ecol.62:399–413
Peeters K, Verleyen E, Hodgson DA, Convey P, Ertz D, et al. 2012. Heterotrophic bacterial diversity inaquatic microbial mat communities from Antarctica. Polar Biol. 35:543–54
Phadtare S. 2004. Recent developments in bacterial cold-shock response. Curr. Issues Mol. Biol. 6:125–36Poland JS, Riddle MJ, Zeeb BA. 2003. Contaminants in the Arctic and the Antarctic: a comparison of sources,
impacts, and remediation options. Polar Rec. 39:369–83Ponder MA, Gilmour SJ, Bergholz PW, Mindock CA, Hollingsworth R, et al. 2005. Characterization of poten-
tial stress responses in ancient Siberian permafrost psychroactive bacteria. FEMS Microbiol. Ecol. 53:103–15Purdy KJ, Nedwell DB, Embley TM. 2003. Analysis of the sulfate-reducing bacterial and methanogenic
archaeal populations in contrasting Antarctic sediments. Appl. Environ. Microbiol. 69:3181–91Rebecchi L, Altiero T, Guidetti R, Cesari M, Bertolani R, et al. 2009. Tardigrade resistance to space effects:
first results of experiments on the LIFE-TARSE mission on FOTON-M3 (September 2007). Astrobiology9:581–91
Reid IN, Sparks WB, Lubnow S, McGrath M, Livio M, et al. 2006. Terrestrial models for extraterrestriallife: methanogens and halophiles at Martian temperatures. Int. J. Astrobiol. 5:89–97
Reid PC, Fischer AC, Lewis-Brown E, Meredith MP, Sparrow M, et al. 2009. Impacts of the oceans onclimate change. Adv. Mar. Biol. 56:1–150
Rodrigues DF, Ivanova N, He Z, Huebner M, Zhou J, Tiedje JM. 2008. Architecture of thermal adaptationin an Exiguobacterium sibiricum strain isolated from 3 million year old permafrost: a genome andtranscriptome approach. BMC Genomics 18:547
Russell NJ. 2008. Membrane components and cold sensing. In Psychrophiles: From Biodiversity to Biotechnology,ed. R Margesin, F Schinner, J-C Marx, C Gerday, pp. 177–90. Berlin: Springer
Sabine CL, Feely RA, Gruber N, Key RM, Lee K, et al. 2004. The oceanic sink for anthropogenic CO2.Science 305:367–71
Saito R, Nakayama A. 2004. Differences in malate dehydrogenases from the obligately piezophilic deep-seabacterium Moritella sp. strain 2D2 and the psychrophilic bacterium Moritella sp. strain 5710. FEMSMicrobiol. Lett. 233:165–72
6.26 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Sarmiento JL, Le Quere C. 1996. Oceanic carbon dioxide uptake in a model of century-scale global warming.Science 274:1346–50
Saunders NFW, Thomas T, Curmi PM, Mattick JS, Kuczek E, et al. 2003. Mechanisms of thermal adaptationrevealed from the genomes of the Antarctic archaea Methanogenium frigidum and Methanococcoides burtonii.Genome Res. 13:1580–88
Sawstrom C, Ganeli W, Laybourn-Parry J, Anesio AM. 2007. High viral infection rates in Antarctic andArctic bacterioplankton. Environ. Microbiol. 9:250–55
Schiaffino MR, Unrein F, Gasol JM, Farias ME, Estevez C, et al. 2009. Comparative analysis of bacterio-plankton assemblages from maritime Antarctic freshwater lakes with contrasting trophic status. PolarBiol. 32:923–36
Sellek GA, Chaudhuri JB. 1999. Biocatalysis in organic media using enzymes from extremophiles. EnzymeMicrob. Technol. 25:471–82
Siddiqui KS, Bokhari SA, Afzal AJ, Singh S. 2004a. A novel thermodynamic relationship based on KramersTheory for studying enzyme kinetics under high viscosity. IUBMB Life 56:403–7
Siddiqui KS, Cavicchioli R. 2006. Cold-adapted enzymes. Annu. Rev. Biochem. 75:403–33Siddiqui KS, Feller G, D’Amico S, Gerday C, Giaquinto L, Cavicchioli R. 2005. The active site is the least stable
structure in the unfolding pathway of a multidomain cold-adapted α-amylase. J. Bacteriol. 187:6197–205Siddiqui KS, Poljak A, Cavicchioli R. 2004b. Improved activity and stability of alkaline phosphatases from
psychrophilic and mesophilic organisms by chemically modifying aliphatic or amino groups usingtetracarboxy-benzophenone derivatives. Cell. Mol. Biol. 50:657–67
Siddiqui KS, Poljak A, Guilhaus M, De Francisci D, Curmi PM, et al. 2006. Role of lysine versus arginine inenzyme cold-adaptation: modifying lysine to homo-arginine stabilizes the cold-adapted α-amylase fromPseudoalteramonas haloplanktis. Proteins 64:486–501
Siddiqui KS, Shemsi AM, Anwar MA, Rashid MH, Rajoka MI. 1999. Partial and complete alteration of sur-face charges of carboxymethylcellulase by chemical modification: thermostabilization in water-miscibleorganic solvent. Enzyme Microbiol. Technol. 24:599–608
Siddiqui KS, Cavicchioli R, Thomas T. 2002. Thermodynamic activation properties of elongation factor 2(EF-2) proteins from psychrotolerant and thermophilic archaea.Extremophiles 6:143–50
Siddiqui KS, Thomas T, eds. 2008. Protein Adaptation in Extremophiles. New York: Nova Sci.Siegert MJ, Ellis-Evans CJ, Tranter M, Mayer C, Petit J-R, et al. 2001. Physical, chemical and biological
processes in Lake Vostok and other Antarctic subglacial lakes. Nature 414:603–9Simon C, Wiezer A, Strittmatter AW, Daniel R. 2009. Phylogenetic diversity and metabolic potential
revealed in a glacier ice metagenome. Appl. Environ. Microbiol. 75:7519–26Snider MJ, Lazarevic D, Wolfenden R. 2002. Catalysis by entropic effects: the action of cytidine deaminase
on 5,6-dihydrocytidine. Biochemistry 41:3925–30Somero GN. 1995. Proteins and temperature. Annu. Rev. Physiol. 57:43–68Somero GN. 2003. Protein adaptations to temperature and pressure: complementary roles of adaptive
changes in amino acid sequence and internal milieu. Comp. Biochem. Physiol. B 136:577–91Somme L, Meier T. 1995. Cold tolerance in Tardigrada from Dronning Maud Land, Antarctica. Polar Biol.
15:221–24Srimathi S, Jayaraman G, Feller G, Danielsson B, Narayanan PR. 2007. Intrinsic halotolerance of the
psychrophilic α-amylase from Pseudoalteromonas haloplanktis. Extremophiles 11:505–15Steig EJ, Schneider DP, Rutherford SD, Mann ME, Comiso JC, Shindell DT. 2009. Warming of the
Antarctic ice-sheet surface since the 1957 International Geophysical Year. Nature 457:459–62Sutherland DL. 2009. Microbial mat communities in response to recent changes in the physiochemical
environment of the meltwater ponds on the McMurdo Ice Shelf, Antarctica. Polar Biol. 32:1023–32Sutherland DL, Hawes I. 2009. Annual growth layers as proxies of past growth conditions for benthic
microbial mats in a perennially ice-covered Antarctic Lake. FEMS Microbiol. Ecol. 67:279–92Takai K, Nakamura K, Toki T, Tsunogai U, Miyazaki M, et al. 2008. Cell proliferation at 122◦C and
isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation.Proc. Natl. Acad. Sci. USA 105:10949–54
Takami H, Inoue A, Fuji F, Horikoshi K. 1997. Microbial flora in the deepest sea mud of the MarianaTrench. FEMS Microbiol. Lett. 152:279–85
www.annualreviews.org • Psychrophiles 6.27
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Tanghe A, Van Dijck P, Thevelein JM. 2003. Determinants of freeze tolerance in microorganisms,physiological importance, and biotechnological applications. Adv. Appl. Microbiol. 53:129–75
Taton A, Grubisic S, Balthasart P, Hodgson DA, Laybourn-Parry J. 2006. Biogeographical distribution andecological ranges of benthis cyanobacteria in East Antarctic Lakes. FEMS Microbiol. Ecol. 57:272–89
Taton A, Grubisic S, Brambilla E, De Wit R, Wilmotte A. 2003. Cyanobacterial diversity in natural andartificial microbial mats of Lake Fryxell (McMurdo Dry Valleys, Antarctica): a morphological andmolecular approach. Appl. Environ. Microbiol. 69:5157–69
Thaler TL, Gibbs PR, Trebino RP, Bommarius AS. 2006. Search for extraterrestrial life using chiralmolecules: mandelate racemase as a test case. Astrobiology 6:901–10
Thomalla SJ, Waldron HN, Lucas MI, Read JF, Ansorge IJ, Pakhomov E. 2011. Phytoplankton distributionand nitrogen dynamics in the Southwest Indian subtropical gyre and Southern Ocean waters. Ocean Sci.7:113–27
Thomas DN, Dieckmann GS. 2002. Antarctic sea ice: a habitat for extremophiles. Science 295:641–44Thomas T, Kumar N, Cavicchioli R. 2001. Effects of ribosomes and intracellular solutes on activities
and stabilities of elongation factor 2 proteins from psychrotolerant and thermophilic methanogens. J.Bacteriol. 183:1974–82
Trent JD. 2000. Extremophiles in astrobiology: per Ardua ad Astra. Gravit. Space Biol. Bull. 13:5–11Unrein F, Izaguirre I, Massana R, Balague V, Gasol JM. 2005. Nanoplankton assemblages in maritime
Antarctic lakes: characterization and molecular fingerprinting comparison. Aquat. Microb. Ecol. 40:269–82Van Trappen S, Mergaert J, Van Eygen S, Dawyndt P, Cnockaert MC, Swings J. 2002. Diversity of 746 het-
erotrophic bacteria isolated from microbial mats from ten Antarctic Lakes. Syst. Appl. Microbiol. 25:603–10Varin T, Lovejoy C, Jungblut AD, Vincent WF, Corbeil J. 2012. Metagenomic analysis of stress genes in
microbial mat communities from Antarctica and the high Artic. Appl. Environ. Microbiol. 78:549–59Verleyen E, Sabbe K, Hodgson DA, Grubisic S, Tato A, et al. 2010. Structuring effects of climate-related
environmental factors on Antarctic microbial mat communities. Aquat. Microb. Ecol. 59:11–24Villaescusa JA, Casamayor EO, Rochera C, Velazquez D, Chicote A, et al. 2010. A close link between bacterial
community composition and environmental heterogeneity in maritime Antarctic lakes. Int. Microbiol.13:67–77
Vincent WF. 2000. Cyanobacterial dominance in the polar regions. In The Ecology of Cyanobacteria: TheirDiversity in Time and Space, ed. BA Whitton, M Potts, pp. 321–40. Dordrecht: Kluwer Acad.
Voytek MA, Priscu JC, Ward BB. 1999. The distribution and relative abundance of ammonia-oxidizingbacteria in the lakes of the McMurdo Dry Valley, Antarctica. Hydrobiologia 401:113–30
Weedon JT, Aerts R, Kowalchuk GA, van Bodegom PM. 2011. Enzymology under global change: organicnitrogen turnover in alpine and sub-arctic soils. Biochem. Soc. Trans. 39:309–14
Whitehouse MJ, Meredith MP, Rothery P, Atkinson A, Ward P, Korb RE. 2008. Rapid warming of theocean around South Georgia, Southern Ocean, during the 20th century: forcings, characteristics andimplications for lower tropic levels. Deep-Sea Res. I 55:1218–28
Wignall PB, Twitchett RJ. 1996. Oceanic anoxia and the end Permian mass extinction. Science 272:1155–58Wilkins D, Lauro FM, Williams TJ, DeMaere MZ, Brown MV, et al. 2012a. Biogeographic partitioning
of Southern Ocean microorganisms revealed by metagenomics. Environ. Microbiol. doi: 10.1111/1462-2920.12035
Wilkins D, Yau S, Williams TJ, Allen MA, Brown MV, et al. 2012b. Key microbial drivers in Antarcticaquatic environments. FEMS Microbiol. Rev. doi: 10.1111/1574-6976.12007
Williams TJ, Burg DW, Ertan H, Raftery MJ, Poljak A, et al. 2010a. Global proteomic analysis of theinsoluble, soluble, and supernatant fractions of the psychrophilic archaeon Methanococcoides burtonii. PartII: the effect of different methylated growth substrates. J. Proteome Res. 9:653–63
Williams TJ, Burg DW, Raftery MJ, Poljak A, Guilhaus M, et al. 2010b. Global proteomic analysis of theinsoluble, soluble, and supernatant fractions of the psychrophilic archaeon Methanococcoides burtonii. PartI: the effect of growth temperature. J. Proteome Res. 9:640–52
Williams TJ, Lauro FM, Ertan H, Burg DW, Poljak A, et al. 2011. Defining the response of a microorganismto temperatures that span its complete growth temperature range (−2◦C to 28◦C) using multiplexquantitative proteomics. Environ. Microbiol. 13:2186–203
6.28 Siddiqui et al.
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.

EA41CH06-Cavicchioli ARI 7 February 2013 14:43
Williams TJ, Long E, Evans F, DeMaere MZ, Lauro FM, et al. 2012a. A metaproteomic assessment of winterand summer bacterioplankton from Antarctic Peninsula coastal surface waters. ISME J. 6:1883–900
Williams TJ, Wilkins D, Long E, Evans F, DeMaere MZ, et al. 2012b. The role of planktonic Flavobac-teria in processing algal organic matter in coastal East Antarctica revealed using metagenomics andmetaproteomics. Environ. Microbiol. doi: 10.1111/1462-2920.12017
Wingham DJ, Siegert MJ, Shepherd A, Muir AS. 2006. Rapid discharge connects Antarctic subglacial lakes.Nature 440:1033–36
Wolfenden R. 2011. Benchmark reaction rates, the stability of biological molecules in water, and the evolutionof catalytic power in enzymes. Annu. Rev. Biochem. 80:645–67
Wolfenden R, Snider MJ. 2001. The depth of chemical time and the power of enzymes as catalysts. Acc.Chem. Res. 34:938–45
Xu Y, Feller G, Gerday C, Glansdorff N. 2003. Moritella cold-active dihydrofolate reductase: Are therenatural limits to optimization of catalytic efficiency at low temperature? J. Bacteriol. 185:5519–26
Yan BQ, Chen XL, Hou XY, He H, Zhou BC, et al. 2009. Molecular analysis of the gene encodinga cold-adapted halophilic subtilase from deep-sea psychrotolerant bacterium Pseudoalteromonas sp.SM9913: cloning, expression, characterization and function analysis of the C-terminal PPC domains.Extremophiles 13:725–33
Yau S, Lauro FM, DeMaere MZ, Brown MV, Thomas T, et al. 2011. Virophage control of Antarctic algalhost-virus dynamics. Proc. Natl. Acad. Sci. USA 108:6163–68
Zhao JS, Deng Y, Manno D, Hawari J. 2010. Shewanella spp. genomic evolution for a cold marine lifestyleand in-situ explosive biodegradation. PLoS ONE 5:e9109
www.annualreviews.org • Psychrophiles 6.29
Changes may still occur before final publication online and in print
Ann
u. R
ev. E
arth
Pla
net.
Sci.
2013
.41.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f N
ew S
outh
Wal
es o
n 02
/24/
13. F
or p
erso
nal u
se o
nly.