growth and physiological features of cyanobacterium anabaena sp. strain pcc 7120 in a...
TRANSCRIPT

BIOTECHNOLOGY AND BIOENGINEERING Chinese Journal of Chemical Engineering, 19(1) 108 115 (2011)
Growth and Physiological Features of Cyanobacterium Anabaena sp. Strain PCC 7120 in a Glucose-Mixotrophic Culture*
YU Guoce ( )1,2,**, SHI Dingji ( )3, CAI Zhaoling ( )2, CONG Wei ( )2
and OUYANG Fan ( )2
1 Institute of Nuclear and New Energy Technology, Tsinghua University, Beijing 100084, China 2 State Key Laboratory of Biochemical Engineering, Institute of Process Engineering, Chinese Academy of Sciences,
Beijing 100080, China 3 Research Center of Photosynthesis, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China
Abstract Mixotrophic growth is one potential mode for mass culture of microalgae and cyanobacteria particularly suitable for the production of high value bioactive compounds and fine chemicals. The typical heterocystous cyanobacterium Anabaena sp. PCC 7120 was grown in the presence of exogenous glucose in light. Glucose im-proved the cell growth evidently, the maximal specific growth rate under mixotrophic condition (0.38 d 1) being 1.6-fold of that of photoautotrophic growth. Mixotrophy caused a variation in cellular pigment composition, in-creasing the content of chlorophyll a and decreasing the contents of carotenoid and phycobiliprotein relative to chlorophyll a. Fluorescence emission from photosystem II (PSII) relative to photosystem I was enhanced in mixotrophic cells, implying an increased energy distribution in PSII. Glucokinase (EC 2.7.1.2) activity was further induced in the presence of glucose. The mixotrophic culture was scaled up in a 15 L airlift photobioreactor equipped with an inner and an outer light source. A modified Monod model incorporating the specific growth rate and the average light intensity in the reactor was developed to describe cell growth appropriately. The understand-ing of mixotrophic growth and relevant physiological features of Anabaena sp. PCC 7120 would be meaningful for cultivation and exploitation of this important cyanobacterial strain. Keywords cyanobacteria, Anabaena sp. PCC 7120, growth features, mixotrophic culture, photobioreactor
1 INTRODUCTION
Mixotrophic culture is a potential mode for mass production of microalgae and cyanobacteria by using heterotrophic capability of the photosynthetic micro-organisms [1 3]. It is expected that mixotrophic growth can achieve high cell densities and synthesize light-induced products such as photosynthetic pig-ments, and is especially suitable for the production of high value bioactive compounds, fine chemicals and pharmaceuticals. Some microalgae and cyanobacteria, which were regarded as obligate photoautotrophs, can utilize organic carbon compounds for their growth [4 6], and the potential of growth depends on the ge-netic background of the specific organism, the nature of the substrate as well as environmental factors. Much work has been conducted on mixotrophic and heterotrophic growth of the green algae Chlorella [7 10], Haematococcus [11 13], Scenedesmus [14, 15], and Platymonas [16]. These work revealed the charac-teristics of mixotrophic growth and the advantage of mixotrophic growth over photoautotrophic growth for biomass production as well as the formation of some specific products in some sense.
Though some strains of cyanobacteria can use various sources of fixed carbon [17, 18], studies on mixotrophic and heterotrophic growth of cyanobacte-ria are limited and little information is available re-garding the physiological features of cyanobacterial
mixotrophy. The existing work has been focused on mixotrophic culture of Spirulina platensis [1, 3, 19 21], with few other cyanobacterial strains studied [22, 23]. Mixotrophic growth of filamentous heterocystous cyanobacteria such as Anabaena, one important cate-gory of cyanobacteria, has rarely been reported [24 26]. It was reported that the production of biomass and photosynthetic pigments during mixotrophic growth of S. platensis was increased by 1.5- to 2.0-fold com-pared with that in photoautotrophic culture [1], and a cell density as high as 10.24 g·L 1 was achieved in fed-batch mixotrophic culture of S. platensis [3]. Dur-ing mixotrophic culture photosynthetic cells utilize light and organic carbon concurrently as energy sources, and CO2 and the organic substrate as carbon sources. The simultaneous involvement of photoauto-trophy and heterotrophy may give rise to a relatively more complicated response of cellular physiology. While Marquez et al. [19] suggested the independence of photosynthetic activity and heterotrophic metabo-lism of S. platensis in mixotrophic culture based on the growth performance in different nutritional modes, it was demonstrated that mixotrophic growth had a significant effect on photosynthetic activity of cyano-bacteria [26 28]. A deep understanding of growth characteristics and intrinsic physiological features of cyanobacteria in mixotrophic culture would be con-structive for the regulation and optimization of the culture technology.
Received 2010-04-06, accepted 2010-09-18.
* Supported by a grant from the State Key Laboratory of Biochemical Engineering, Institute of Process Engineering, Chinese Academy of Sciences.
** To whom correspondence should be addressed. E-mail: [email protected]

Chin. J. Chem. Eng., Vol. 19, No. 1, February 2011 109
Anabaena sp. strain PCC 7120 is a typical fila-mentous heterocystous cyanobacterium, which has been adopted as a model species for investigation of cyanobacterial physiology and molecular biology [29, 30], and its entire genome sequence has been determined [31]. Anabaena sp. PCC 7120 can be cultured for the production of a variety of fine chemicals and bioactive compounds, such as sulfolipid [32], pigments and exopolysaccharides [33]. Furthermore, since Anabaena sp. PCC 7120 is used as a conventional host strain in cyanobacterial genetic engineering, it is very promis-ing to produce high value transgenic proteins through cultivation of this cyanobacterial strain [34, 35]. So far, few studies have been reported on the culture of Ana-baena sp. PCC 7120 [32, 33], and the potential of mixotrophic growth of the cyanobacterium has rarely been explored to our knowledge.
In our preliminary work Anabaena sp. PCC 7120 was found to be able to grow on glucose in light and not in darkness, indicating its mixotrophic but not heterotrophic capability. The current work presents some characteristic features of mixotrophic growth of Anabaena sp. PCC 7120 with glucose as the sole ex-ogenous carbon substrate. Scale up of mixotrophic culture in a new type airlift photobioreactor and rele-vant kinetic analysis of mixotrophic growth are also carried out.
2 MATERIALS AND METHODS
2.1 Strain and medium
The cyanobacterium Anabaena sp. strain PCC 7120 was obtained from the Research Center of Pho-tosynthesis, Institute of Botany, Chinese Academy of Sciences, Beijing. The basal medium BG-11 [36] was adopted throughout the study, in which sodium chloride was substituted for sodium nitrate unless otherwise indicated and the buffer Tris-HCl (pH 7.5) was used. Glucose was added to give a required initial concen-tration as an exogenous carbon source in the medium.
2.2 Shake flask culture
Anabaena sp. PCC 7120 was maintained on plates of BG-11 medium, solidified with mass con-centration of 1.5% agar powder. Cells were subcul-tured in a 50 ml Erlenmeyer flask containing 20 ml BG-11 medium at 30 C, 130 r·min 1 and a light in-tensity of 160 E·m 2·s 1, and then transferred to a 250 ml Erlenmeyer flask containing 130 ml medium. After 5-day cultivation, 20 ml cell suspension was inoculated to 250 ml Erlenmeyer flasks, each contain-ing 130 ml medium with glucose concentration ar-ranged as indicated. The irradiance was provided with five 18 W fluorescent lamps (PHILIPS TLD 18W/54). Continuous illumination with a fixed intensity of 160
E·m 2·s 1 was guaranteed throughout all cultures. A sample of 5 ml was taken aseptically at a fixed time
every three days. Experiments were performed in du-plicate for each case to be examined. Results were expressed as the mean values, and relative standard deviations were less than 10%.
2.3 Photobioreactor culture
The photobioreactor system consisted of a 1.8 L bubble column photobioreactor (BCR, 1.5 L working volume) and a 15 L airlift photobioreactor (ALR, 11 L working volume) (Fig. 1). With a ratio of illuminating area to volume of 33.3 m 1, the BCR was irradiated with four 18 W fluorescent lamps giving the maxi-mum surface light intensity of 624 E·m 2·s 1 and the minimum of 110 E·m 2·s 1. The ALR (reactor height 1 m, draft tube height 0.6 m, ratio of the cross-sectional area of the riser to the downcomer 0.75) was charac-terized by the outer and inner light source, possessing a ratio of illuminating area to volume of 33.2 m 1. Sixteen 30 W fluorescent lamps (PHILIPS TLD 30W/54) were used as the outer light source, provid-ing an average light intensity of 339.1 E·m 2·s 1 on the outer surface, and one 30 W fluorescent lamp as the inner light source, giving an average light intensity of 228.5 E·m 2·s 1 on the inner surface.
The BCR was inoculated with pure culture grown in shake flasks and operated at 30 C and air aeration of 0.178 vvm. A four-day BCR photoautotrophic cul-ture was pressed into the ALR aseptically and adopted as the inoculum (13%) for the ALR cultivation. The ALR was operated at 30 C and an aeration rate (air containing 3% CO2) of 0.227 vvm. Nitrogenous BG-11 medium was used in photobioreactor cultures and a glucose concentration of 3 g·L 1 was used in mixotrophic culture in the ALR. The BCR was steril-ized in an autoclave and the ALR by steam in situ prior to cultivation.
2.4 Analytical methods
Cell concentration was determined by measuring the absorbance at 750 nm, which displayed linear rela-tionship with dry cell mass within a scope of OD750 from 0 to 0.6. The chlorophyll (Chl) concentration was calculated according to the following formula [37], Chl ( g·ml 1) 14.96 (OD678 OD750) 0.616 (OD720 OD750), where OD678 and OD720 were obtained at the same time as OD750.
Samples were centrifuged at 4000 r·min 1 for 20 min with the harvested supernatant analyzed for glu-cose. Glucose was determined by using the Fehling method or a glucose analyzer (YSI MODEL 2700 SELECT, Yellow Springs Instruments, USA).
Light intensity (photon flux density) was meas-ured by using a photosynthetically active radiometer (FGH-1, Factory of Photoelectric Instruments of Bei-jing Normal University, Beijing, China).
Absorption spectra at room temperature (25 C) were measured by using a dual wavelength double-beam

Chin. J. Chem. Eng., Vol. 19, No. 1, February 2011 110
recording spectrophotometer (UV3000, Shimadzu, Japan). Fluorescence emission spectra at 77 K were measured by using a fluorescence spectrophotometer (F-4500, Hitachi, Japan). Two exciting light wave-lengths of 436 nm and 580 nm were adopted.
Glucokinase (EC 2.7.1.2) activity was measured according to Slein et al. [38] and Pearce and Carr [39]. Protein concentration was determined by the modified Lowry method [40]. Exponentially growing cells were washed with deionized water, suspended in 0.1 mol·L 1 potassium phosphate buffer (pH 7.0) and broken by ultrasonic distintegration in ice bath. The resultant suspension was centrifuged at 9000 r·min 1 for 10 min at 2 C yielding the supernatant for enzyme and protein assay. Analysis was done in triplicate, and results were expressed as means of three determina-tions standard deviations.
The attenuation of light in cultures was deter-mined as described by Hirata et al. [41] with an inci-dent light intensity of 234.0 E·m 2·s 1.
2.5 Average light intensity in the airlift photobio-reactor
The following assumptions are made for the es-timation of the average light intensity in the ALR: (1) the incident light intensity on the reactor surface is uniform; (2) there is no axial distribution of light in-tensity in the reactor; and (3) the attenuation of light obeys the Lambert-Beer law. Light intensity can thus be taken as a function of cell density and radial posi-tion in the photobioreactor.
According to the Lambert-Beer law, the light in-
tensity yielded by one beam from the outer light source at radial position r can be expressed as
out ( , )out out( , ) e p rI r I (1)
where Iout is the light intensity at outer surface of re-actor, is the absorption coefficient, and pout( ,r) is the length of light path along which the beam goes from outer surface of reactor to radial position r at an angle , i.e. 2 2 2 0.5
out 2( , ) cos ( sin )p r r R r , where R2 is the reactor radius. The whole outer light source yields the light intensity at radial position r
1out outarcsin( / )
( ) 2 ( , )dR r
I r I r (2)
where R1 is the radius of the tube containing the inner light source.
Similarly, the whole inner light source gives the light intensity at radial position r
1arcsin( / )in in0
( ) 2 ( , )dR r
I r I r (3)
where in ( , )in in( , ) e p rI r I , in which Iin is the light
intensity at inner surface of reactor and in ( , )p r 2 2 2 0.51cos ( sin )r R r .
Thus, the light intensity at radial position r is
out in( ) ( ) ( )I r I r I r (4)
The average light intensity in the ALR can be obtained as follows.
2
21
2 1
1
av2 1
( )d 1 ( )dd
R
RRR R
R
I r rI I r r
R Rr (5)
Figure 1 Schematic diagram of the photobioreactor system
Chin. J. Chem. Eng., Vol. 19, No. 1, February 2011 111
The average light intensities at different cell den-sities can be derived through numerical integration by using the Mathematica software.
3 RESULTS AND DISCUSSION
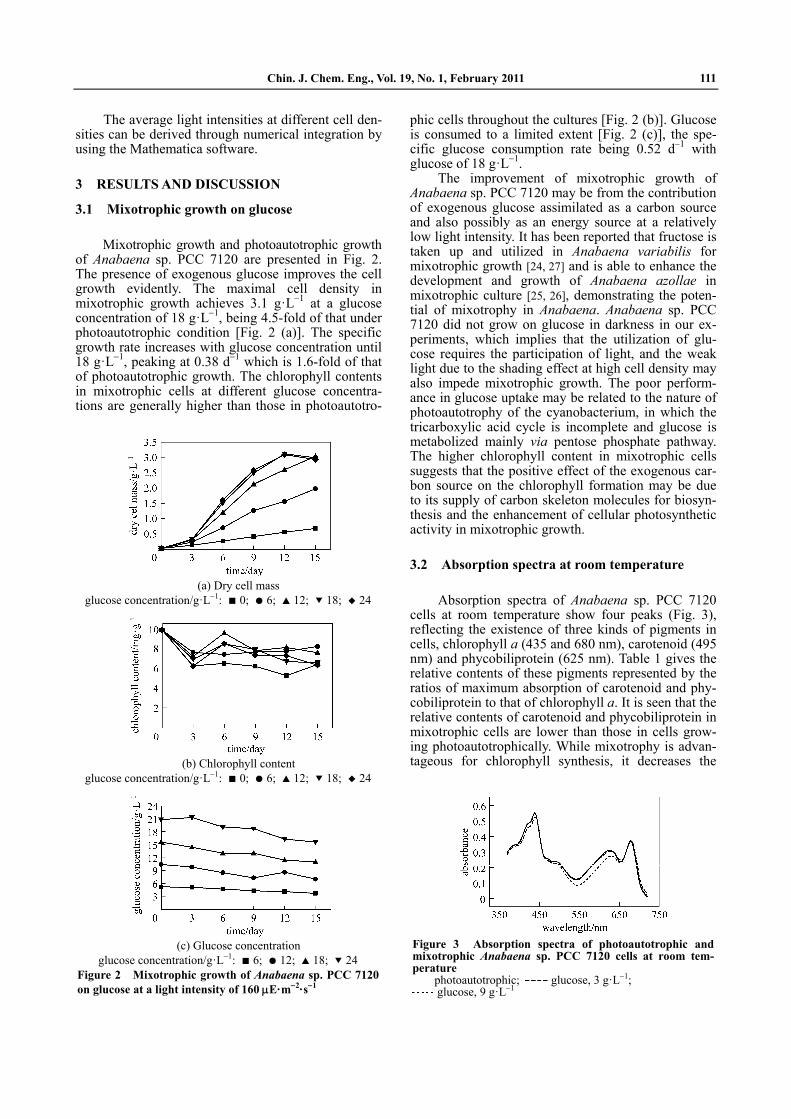
3.1 Mixotrophic growth on glucose
Mixotrophic growth and photoautotrophic growth of Anabaena sp. PCC 7120 are presented in Fig. 2. The presence of exogenous glucose improves the cell growth evidently. The maximal cell density in mixotrophic growth achieves 3.1 g·L 1 at a glucose concentration of 18 g·L 1, being 4.5-fold of that under photoautotrophic condition [Fig. 2 (a)]. The specific growth rate increases with glucose concentration until 18 g·L 1, peaking at 0.38 d 1 which is 1.6-fold of that of photoautotrophic growth. The chlorophyll contents in mixotrophic cells at different glucose concentra-tions are generally higher than those in photoautotro-
phic cells throughout the cultures [Fig. 2 (b)]. Glucose is consumed to a limited extent [Fig. 2 (c)], the spe-cific glucose consumption rate being 0.52 d 1 with glucose of 18 g·L 1.
The improvement of mixotrophic growth of Anabaena sp. PCC 7120 may be from the contribution of exogenous glucose assimilated as a carbon source and also possibly as an energy source at a relatively low light intensity. It has been reported that fructose is taken up and utilized in Anabaena variabilis for mixotrophic growth [24, 27] and is able to enhance the development and growth of Anabaena azollae in mixotrophic culture [25, 26], demonstrating the poten-tial of mixotrophy in Anabaena. Anabaena sp. PCC 7120 did not grow on glucose in darkness in our ex-periments, which implies that the utilization of glu-cose requires the participation of light, and the weak light due to the shading effect at high cell density may also impede mixotrophic growth. The poor perform-ance in glucose uptake may be related to the nature of photoautotrophy of the cyanobacterium, in which the tricarboxylic acid cycle is incomplete and glucose is metabolized mainly via pentose phosphate pathway. The higher chlorophyll content in mixotrophic cells suggests that the positive effect of the exogenous car-bon source on the chlorophyll formation may be due to its supply of carbon skeleton molecules for biosyn-thesis and the enhancement of cellular photosynthetic activity in mixotrophic growth.
3.2 Absorption spectra at room temperature
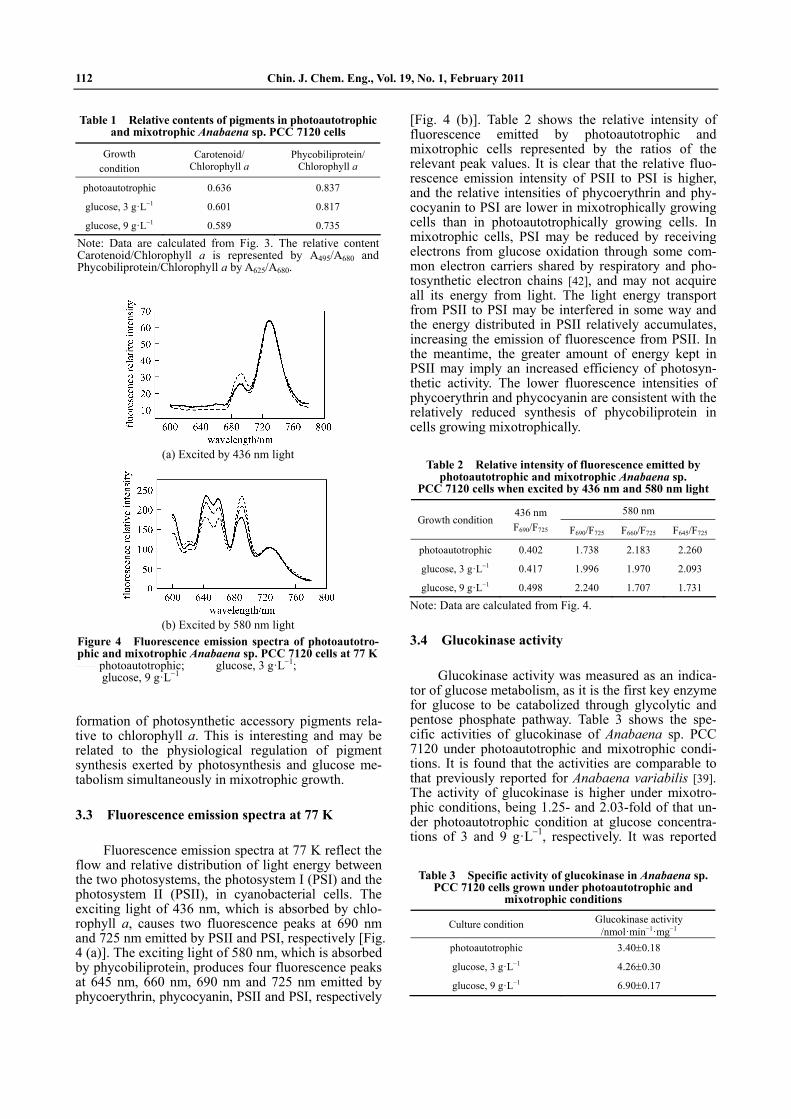
Absorption spectra of Anabaena sp. PCC 7120 cells at room temperature show four peaks (Fig. 3), reflecting the existence of three kinds of pigments in cells, chlorophyll a (435 and 680 nm), carotenoid (495 nm) and phycobiliprotein (625 nm). Table 1 gives the relative contents of these pigments represented by the ratios of maximum absorption of carotenoid and phy-cobiliprotein to that of chlorophyll a. It is seen that the relative contents of carotenoid and phycobiliprotein in mixotrophic cells are lower than those in cells grow-ing photoautotrophically. While mixotrophy is advan-tageous for chlorophyll synthesis, it decreases the
(a) Dry cell mass
glucose concentration/g·L 1: 0; 6; 12; 18; 24
(b) Chlorophyll content
glucose concentration/g·L 1: 0; 6; 12; 18; 24
(c) Glucose concentration
glucose concentration/g·L 1: 6; 12; 18; 24Figure 2 Mixotrophic growth of Anabaena sp. PCC 7120 on glucose at a light intensity of 160 E·m 2·s 1
Figure 3 Absorption spectra of photoautotrophic and mixotrophic Anabaena sp. PCC 7120 cells at room tem-perature
photoautotrophic; glucose, 3 g·L 1; glucose, 9 g·L 1
Chin. J. Chem. Eng., Vol. 19, No. 1, February 2011 112
formation of photosynthetic accessory pigments rela-tive to chlorophyll a. This is interesting and may be related to the physiological regulation of pigment synthesis exerted by photosynthesis and glucose me-tabolism simultaneously in mixotrophic growth.
3.3 Fluorescence emission spectra at 77 K
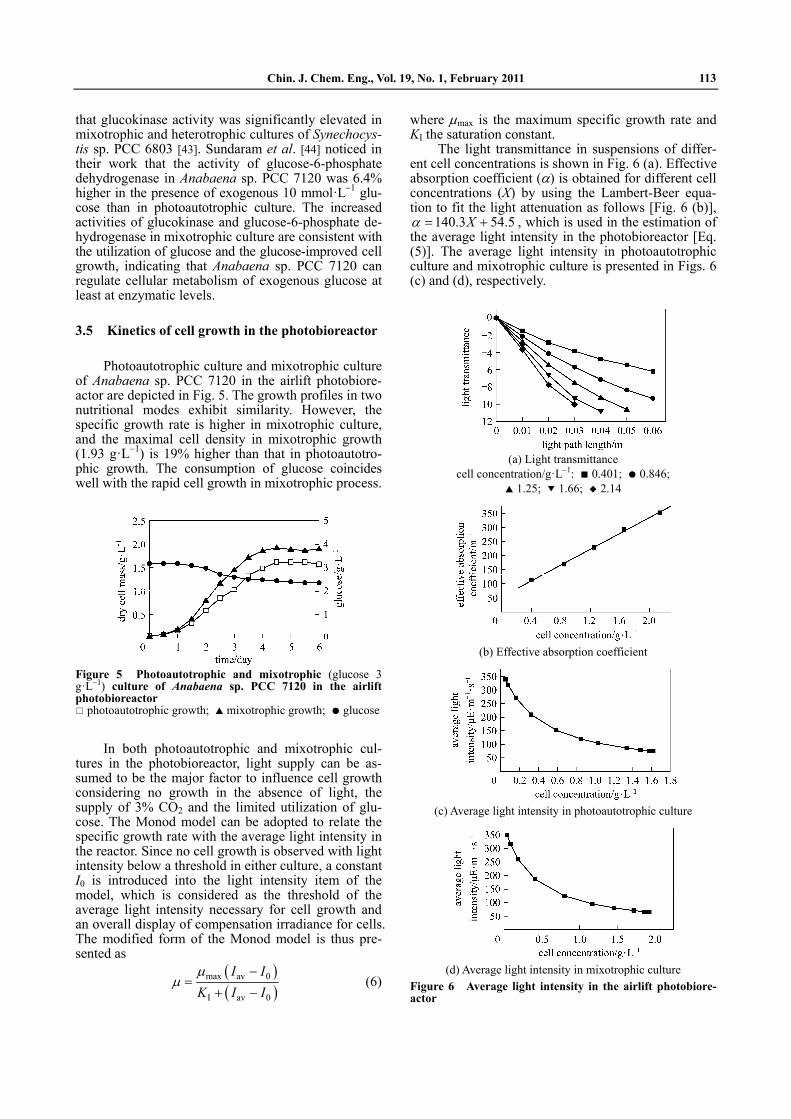
Fluorescence emission spectra at 77 K reflect the flow and relative distribution of light energy between the two photosystems, the photosystem I (PSI) and the photosystem II (PSII), in cyanobacterial cells. The exciting light of 436 nm, which is absorbed by chlo-rophyll a, causes two fluorescence peaks at 690 nm and 725 nm emitted by PSII and PSI, respectively [Fig. 4 (a)]. The exciting light of 580 nm, which is absorbed by phycobiliprotein, produces four fluorescence peaks at 645 nm, 660 nm, 690 nm and 725 nm emitted by phycoerythrin, phycocyanin, PSII and PSI, respectively
[Fig. 4 (b)]. Table 2 shows the relative intensity of fluorescence emitted by photoautotrophic and mixotrophic cells represented by the ratios of the relevant peak values. It is clear that the relative fluo-rescence emission intensity of PSII to PSI is higher, and the relative intensities of phycoerythrin and phy-cocyanin to PSI are lower in mixotrophically growing cells than in photoautotrophically growing cells. In mixotrophic cells, PSI may be reduced by receiving electrons from glucose oxidation through some com-mon electron carriers shared by respiratory and pho-tosynthetic electron chains [42], and may not acquire all its energy from light. The light energy transport from PSII to PSI may be interfered in some way and the energy distributed in PSII relatively accumulates, increasing the emission of fluorescence from PSII. In the meantime, the greater amount of energy kept in PSII may imply an increased efficiency of photosyn-thetic activity. The lower fluorescence intensities of phycoerythrin and phycocyanin are consistent with the relatively reduced synthesis of phycobiliprotein in cells growing mixotrophically.
Table 2 Relative intensity of fluorescence emitted by photoautotrophic and mixotrophic Anabaena sp.
PCC 7120 cells when excited by 436 nm and 580 nm light
580 nm Growth condition
436 nm F690/F725 F690/F725 F660/F725 F645/F725
photoautotrophic 0.402 1.738 2.183 2.260
glucose, 3 g·L 1 0.417 1.996 1.970 2.093
glucose, 9 g·L 1 0.498 2.240 1.707 1.731
Note: Data are calculated from Fig. 4.
3.4 Glucokinase activity
Glucokinase activity was measured as an indica-tor of glucose metabolism, as it is the first key enzyme for glucose to be catabolized through glycolytic and pentose phosphate pathway. Table 3 shows the spe-cific activities of glucokinase of Anabaena sp. PCC 7120 under photoautotrophic and mixotrophic condi-tions. It is found that the activities are comparable to that previously reported for Anabaena variabilis [39]. The activity of glucokinase is higher under mixotro-phic conditions, being 1.25- and 2.03-fold of that un-der photoautotrophic condition at glucose concentra-tions of 3 and 9 g·L 1, respectively. It was reported
Table 1 Relative contents of pigments in photoautotrophicand mixotrophic Anabaena sp. PCC 7120 cells
Growth condition
Carotenoid/ Chlorophyll a
Phycobiliprotein/ Chlorophyll a
photoautotrophic 0.636 0.837
glucose, 3 g·L 1 0.601 0.817
glucose, 9 g·L 1 0.589 0.735
Note: Data are calculated from Fig. 3. The relative content Carotenoid/Chlorophyll a is represented by A495/A680 andPhycobiliprotein/Chlorophyll a by A625/A680.
(a) Excited by 436 nm light
(b) Excited by 580 nm light
Figure 4 Fluorescence emission spectra of photoautotro-phic and mixotrophic Anabaena sp. PCC 7120 cells at 77 K
photoautotrophic; glucose, 3 g·L 1; glucose, 9 g·L 1
Table 3 Specific activity of glucokinase in Anabaena sp. PCC 7120 cells grown under photoautotrophic and
mixotrophic conditions
Culture condition Glucokinase activity /nmol·min 1·mg 1
photoautotrophic 3.40 0.18 glucose, 3 g·L 1 4.26 0.30 glucose, 9 g·L 1 6.90 0.17
Chin. J. Chem. Eng., Vol. 19, No. 1, February 2011 113
that glucokinase activity was significantly elevated in mixotrophic and heterotrophic cultures of Synechocys-tis sp. PCC 6803 [43]. Sundaram et al. [44] noticed in their work that the activity of glucose-6-phosphate dehydrogenase in Anabaena sp. PCC 7120 was 6.4% higher in the presence of exogenous 10 mmol·L 1 glu-cose than in photoautotrophic culture. The increased activities of glucokinase and glucose-6-phosphate de-hydrogenase in mixotrophic culture are consistent with the utilization of glucose and the glucose-improved cell growth, indicating that Anabaena sp. PCC 7120 can regulate cellular metabolism of exogenous glucose at least at enzymatic levels.
3.5 Kinetics of cell growth in the photobioreactor
Photoautotrophic culture and mixotrophic culture of Anabaena sp. PCC 7120 in the airlift photobiore-actor are depicted in Fig. 5. The growth profiles in two nutritional modes exhibit similarity. However, the specific growth rate is higher in mixotrophic culture, and the maximal cell density in mixotrophic growth (1.93 g·L 1) is 19% higher than that in photoautotro-phic growth. The consumption of glucose coincides well with the rapid cell growth in mixotrophic process.
Figure 5 Photoautotrophic and mixotrophic (glucose 3 g·L 1) culture of Anabaena sp. PCC 7120 in the airlift photobioreactor
photoautotrophic growth; mixotrophic growth; glucose
In both photoautotrophic and mixotrophic cul-tures in the photobioreactor, light supply can be as-sumed to be the major factor to influence cell growth considering no growth in the absence of light, the supply of 3% CO2 and the limited utilization of glu-cose. The Monod model can be adopted to relate the specific growth rate with the average light intensity in the reactor. Since no cell growth is observed with light intensity below a threshold in either culture, a constant I0 is introduced into the light intensity item of the model, which is considered as the threshold of the average light intensity necessary for cell growth and an overall display of compensation irradiance for cells. The modified form of the Monod model is thus pre-sented as
max av 0
I av 0
I IK I I
(6)
where max is the maximum specific growth rate and KI the saturation constant.
The light transmittance in suspensions of differ-ent cell concentrations is shown in Fig. 6 (a). Effective absorption coefficient ( ) is obtained for different cell concentrations (X) by using the Lambert-Beer equa-tion to fit the light attenuation as follows [Fig. 6 (b)],
140.3 54.5X , which is used in the estimation of the average light intensity in the photobioreactor [Eq. (5)]. The average light intensity in photoautotrophic culture and mixotrophic culture is presented in Figs. 6 (c) and (d), respectively.
(a) Light transmittance
cell concentration/g·L 1: 0.401; 0.846; 1.25; 1.66; 2.14
(b) Effective absorption coefficient
(c) Average light intensity in photoautotrophic culture
(d) Average light intensity in mixotrophic culture
Figure 6 Average light intensity in the airlift photobiore-actor

Chin. J. Chem. Eng., Vol. 19, No. 1, February 2011 114
The results by using the modified Monod model to fit photoautotrophic and mixotrophic growth in the photobioreactor (Fig. 5) are shown in Fig. 7. It is seen that the model gives an appropriate description of cell growth. Table 4 lists the estimated parameters of the kinetic model from the data of photoautotrophic and mixotrophic growth. The maximum specific growth rate of mixotrophic growth is greater than that of photoautotrophic growth, and the I0 value and the saturation constant for mixotrophic growth are lower than those for photoautotrophic growth. These results suggest that mixotrophic cells can start to grow at a lower average light intensity and achieve a higher specific growth rate when adopting the same light in-tensity as in photoautotrophic culture. That is, Ana-baena sp. PCC 7120 may depend to a lesser degree on light in mixotrophic growth and can realize a better cell growth compared with photoautotrophic culture due to the participation of glucose and possibly the more efficient use of light.
(a) Photoautotrophic growth
(b) Mixotrophic growth
Figure 7 Photoautotrophic and mixotrophic growth of Anabaena sp. PCC 7120 in the airlift photobioreactor and their fitted results by the modified Monod model
Table 4 Comparison of estimated parameters of the modified Monod model from data of photoautotrophic
and mixotrophic growth of Anabaena sp. PCC 7120 in the airlift photobioreactor
max/d 1 I0/ E·m 2·s 1 KI/ E·m 2·s 1
photoautotrophic growth 1.759 73.04 71.22 mixotrophic growth 2.081 64.84 62.54
4 CONCLUSIONS
This paper reported some characteristic features
of mixotrophic growth and physiology of Anabaena sp. PCC 7120 with glucose as the exogenous carbon substrate. The existence of glucose was able to im-prove cell growth evidently. Mixotrophy modified the composition of cellular photosynthetic pigments and the energy flow between two photosystems. The activ-ity of glucose assimilation was further stimulated in mixotrophic growth. Cell growth in the airlift photo-bioreactor and relevant kinetic analysis revealed that mixotrophic culture can realize a more advantageous growth than photoautotrophic culture for this impor-tant cyanobacterial strain.
ACKNOWLEDGEMENTS
The authors wish to thank Miss Ning Shao and Miss Pengpeng Zhang for their technical assistance in spectra measurement.
REFERENCES
1 Marquez, F.J., Nishio, N., Nagai, S., Sasaki, K., “Enhancement of biomass and pigment production during growth of Spirulina platen-sis in mixotrophic culture”, J. Chem. Technol. Biotechnol., 62, 159 164 (1995).
2 Chen, F., “High cell density culture of microalgae in heterotrophic growth”, Trends Biotechnol., 14, 421 426 (1996).
3 Chen, F., Zhang, Y., “High cell density mixotrophic culture of Spirulina platensis on glucose for phycocyanin production using a fed-batch system”, Enzyme Microb. Technol., 20, 221 224 (1997).
4 Droop, M.R., “Heterotrophy of carbon”, In: Algal Physiology and Biochemistry, University of California Press, Berkeley and Los An-geles, 530 559 (1974).
5 Smith, A.J., “Modes of cyanobacterial carbon metabolism”, In: The Biology of Cyanobacteria, Blackwell Scientific Publications, Oxford, London, Edinburgh, Boston and Melbourne, 47 85 (1982).
6 Tuchman, N.C., “The role of heterotrophy in algae”, In: Algal Ecol-ogy, Academic Press, San Diego, 299 319 (1996).
7 Lee, Y.K., Ding, S.Y., Hoe, C.H., Low, C.S., “Mixotrophic growth of Chlorella sorokiniana in outdoor enclosed photobioreactor”, J. Appl. Phycol., 8, 163 169 (1996).
8 Martinez, M.E., Camacho, F., Jimenez, J.M., Espinola, J.B., “Influ-ence of light intensity on the kinetic and yield parameters of Chlor-ella pyrenoidosa mixotrophic growth”, Process Biochem., 32, 93 98 (1997).
9 Ip, P.F., Wong, K.H., Chen, F., “Enhanced production of astaxanthin by the green microalga Chlorella zofingiensis in mixotrophic cul-ture”, Process Biochem., 39, 1761 1766 (2004).
10 Liang, Y., Sarkany, N., Cui, Y., “Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions”, Biotechnol. Lett., 31, 1043 1049 (2009).
11 Kobayashi, M., Kakizono, T., Yamaguchi, K., Nishio, N., Nagai, S., “Growth and astaxanthin formation of Haematococcus pluvialis in heterotrophic and mixotrophic conditions”, J. Ferment. Bioeng., 74, 17 20 (1992).
12 Zhang, X.W., Gong, X.D., Chen, F., “Kinetic models for astaxanthin production by high cell density mixotrophic culture of the microalga Haematococcus pluvialis”, J. Ind. Microbiol. Biotechnol., 23, 691 696 (1999).
13 Jeon, Y.C., Cho, C.W., Yun, Y.S., “Combined effects of light inten-sity and acetate concentration on the growth of unicellular microalga Haematococcus pluvialis”, Enzyme Microb. Technol., 39, 490 495 (2006).
14 Shamala, T.R., Drawert, F., Leupold, G., “Studies on Scenedesmus acutus growth. I. Effect of autotrophic and mixotrophic conditions on the growth of Scenedesmus acutus”, Biotechnol. Bioeng., 24, 1287 1299 (1982).

Chin. J. Chem. Eng., Vol. 19, No. 1, February 2011 115
15 Combres, C., Laliberte, G., Reyssac, J.S., Delanoue, J., “Effect of acetate on growth and ammonium uptake in the microalga Scenedesmus obliquus”, Physiol. Plant., 91, 729 734 (1994).
16 Xie, J., Zhang, Y., Li, Y., Wang, Y., “Mixotrophic cultivation of Platymonas subcordiformis”, J. Appl. Phycol., 13, 343 347 (2001).
17 Rippka, R., “Photoheterotrophy and chemoheterotrophy among uni-cellular blue-green algae”, Arch. Microbiol., 87, 93 98 (1972).
18 Stal, L.J., Moezelaar, R., “Fermentation in cyanobacteria”, FEMS Microbiol. Rev., 21, 179 211 (1997).
19 Marquez, F.J., Sasaki, K., Kakizono, T., Nishio, N., Nagai, S., “Growth characteristics of Spirulina platensis in mixotrophic and heterotrophic conditions”, J. Ferment. Bioeng., 76, 408 410 (1993).
20 Chen, F., Zhang, Y., Guo, S., “Growth and phycocyanin formation of Spirulina platensis in photoheterotrophic culture”, Biotechnol. Lett., 18, 603 608 (1996).
21 Chen, T., Zheng, W., Yang, F., Bai, Y., Wong, Y.S., “Mixotrophic culture of high selenium-enriched Spirulina platensis on acetate and the enhanced production of photosynthetic pigments”, Enzyme Mi-crob. Technol., 39, 103 107 (2006).
22 Wang, Y., Li, Y., Shi, D., Shen, G., Ru, B., Zhang, S., “Characteris-tics of mixotrophic growth of Synechocystis sp. in an enclosed photobioreactor”, Biotechnol. Lett., 24, 1593 1597 (2002).
23 Yu, H., Jia, S., Dai, Y., “Growth characteristics of the cyanobacte-rium Nostoc flagelliforme in photoautotrophic, mixotrophic and het-erotrophic cultivation”, J. Appl. Phycol., 21, 127 133 (2009).
24 Haury, J.F., Spiller, H., “Fructose uptake and influence on growth of and nitrogen fixation by Anabaena variabilis”, J. Bacteriol., 147, 227 235 (1981).
25 Rozen, A., Arad, H., Schonfeld, M., Tel-Or, E., “Fructose supports glycogen accumulation, heterocysts differentiation, N2 fixation and growth of the isolated cyanobiont Anabaena azollae”, Arch. Micro-biol., 145, 187 190 (1986).
26 Rozen, A., Schonfeld, M., Tel-Or, E., “Fructose-enhanced develop-ment and growth of the N2-fixing cyanobiont Anabaena azollae”, Z. Naturforsch., 43c, 408 412 (1988).
27 Valiente, E.F., Nieva, M., Avendano, M.C., Maeso, E.S., “Uptake and utilization of fructose by Anabaena variabilis ATCC 29413. Ef-fect on respiration and photosynthesis”, Plant Cell Physiol., 33, 307 313 (1992).
28 Vonshak, A., Cheung, S.M., Chen, F., “Mixotrophic growth modifies the response of Spirulina (Arthrospira) platensis (cyanobacteria) cells to light”, J. Phycol., 36, 675 679 (2000).
29 Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M., Stanier, R.Y., “Generic assignments, strain histories and properties of pure cultures of cyanobacteria”, J. Gen. Microbiol., 111, 1 61 (1979).
30 Bryant, D.A., The Molecular Biology of Cyanobacteria, Kluwer Academic Publishers, The Netherlands (1994).
31 Kaneko, T., Nakamura, Y., Wolk, C.P., Kuritz, T., Sasamoto, S., Wa-tanabe, A., Iriguchi, M., Ishikawa, A., Kawashima, K., Kimura, T., Kishida, Y., Kohara, M., Matsumoto, M., Matsuno, A., Muraki, A.,
Nakazaki, N., Shimpo, S., Sugimoto, M., Takazawa, M., Yamada, M., Yasuda, M., Tabata, S., “Complete genomic sequence of the fila-mentous nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120”, DNA Res., 8, 227 253 (2001).
32 Archer, S.D., McDonald, K.A., Jackman, A.P., “Effect of light ir-radiance on the production of sulfolipids from Anabaena 7120 in a fed-batch photobioreactor”, Appl. Biochem. Biotechnol., 67, 139 152 (1997).
33 Morales, E., Rodriguez, M., Garcia, D., Loreto, C., Marco, E., “Ef-fect of pH and CO2 on growth, pigments and exopolysaccharides production from cyanobacteria Anabaena sp. PCC 7120”, Intercien-cia, 27, 373 378 (2002).
34 Ren, L., Shi, D.J., Dai, J.X., Ru, B.G., “Expression of the mouse metallothionein-I gene conferring cadmium resistance in a transgenic cyanobacterium”, FEMS Microbiol. Lett., 158, 127 132 (1998).
35 Liu, F.L., Zhang, H.B., Shi, D.J., Shang, Z.D., Lin, C., Shao, N., Peng, G.H., Zhang, X.Y., Zhang, H.X., Wu, J.Y., Wang, J., Xu, X.D., Jiang, Y.H., Zhong, Z.P., Zhao, S.J., Wu, M., Zeng, C.K., “Construc-tion of shuttle, expression vector of human tumor necrosis factor al-pha (hTNF- ) gene and its expression in a cyanobacterium, Ana-baena sp. PCC7120”, Sci. China (Series C), 42, 25 33 (1999).
36 Castenholz, R.W., “Culturing methods for cyanobacteria”, In: Methods in Enzymology, Vol. 167, Cyanobacteria, Academic Press, San Diego, 68 93 (1988).
37 Williams, J.G.K., “Construction of specific mutations in photosystem II photosynthetic reaction center by genetic engineering methods in Synechocystis 6803”, In: Methods in Enzymology, Vol. 167, Cyano-bacteria, Academic Press, San Diego, 766 778 (1988).
38 Slein, M.W., Cori, G.T., Cori, C.F., “A comparative study of hexokinase from yeast and animal tissues”, J. Biol. Chem., 186, 763 780 (1950).
39 Pearce, J., Carr, N.G., “The incorporation and metabolism of glucose by Anabaena variabilis”, J. Gen. Microbiol., 54, 451 462 (1969).
40 Markwell, M.A.K., Haas, S.M., Bieber, L.L., Tolbert, N.E., “A modification of the Lowry procedure to simplify protein determina-tion in membrane and lipoprotein samples”, Anal. Biochem., 87, 206 210 (1978).
41 Hirata, S., Taya, M., Tone, S., “Characterization of Chlorella cell cultures in batch and continuous operations under a photoautotrophic condition”, J. Chem. Eng. Japan, 29, 953 959 (1996).
42 Scherer, S., “Do photosynthetic and respiratory electron transport chains share redox proteins”, Trends Biochem. Sci., 15, 458 462 (1990).
43 Knowles, V.L., Plaxton, W.C., “From genome to enzyme: analysis of key glycolytic and oxidative pentose-phosphate pathway enzymes in the cyanobacterium Synechocystis sp. PCC 6803”, Plant Cell Physiol., 44, 758 763 (2003).
44 Sundaram, S., Karakaya, H., Scanlan, D.J., Mann, N.H., “Multiple oligomeric forms of glucose-6-phosphate dehydrogenase in cyano-bacteria and the role of OpcA in the assembly process”, Microbiol-ogy, 144, 1549 1556 (1998).