title 苔類ゼニゴケにおけるフィトクロムシグナル伝達機構の … ·...
TRANSCRIPT

Title 苔類ゼニゴケにおけるフィトクロムシグナル伝達機構の解析( Dissertation_全文 )
Author(s) 井上, 佳祐
Citation Kyoto University (京都大学)
Issue Date 2016-09-23
URL https://doi.org/10.14989/doctor.k20035
Right
Type Thesis or Dissertation
Textversion ETD
Kyoto University

苔類ゼニゴケにおけるフィトクロム
シグナル伝達機構の解析
井上 佳祐

1
目次
要旨 2
略語表 3
序論 5
結果 13
考察 35
材料と方法 39
文献 45
謝辞 54

2
要旨
フィトクロムは赤色光および遠赤色光を受容する光受容体であり、植物の成長や発生の
多くの面で重要な役割をもつ。シロイヌナズナを用いた解析から、フィトクロムは主に核内
で機能し、赤色光シグナルの抑制因子として働く PHYTOCHROME INTERACTING FACTOR
(PIF) という bHLH型転写因子の機能を阻害することで遺伝子発現を制御することが知られ
ている。これまでに、赤色光/遠赤色光可逆的な応答やフィトクロム遺伝子の存在は多くの
植物種で報告されているが、シロイヌナズナ以外の植物種におけるフィトクロムシグナル
伝達の分子機構に関してはほとんど知見がない。そこで本研究では基部陸上植物である苔
類ゼニゴケを用いて、フィトクロム自身の性質の解析とともに、そのシグナル伝達機構につ
いて解析を行った。
まず、ゼニゴケにおけるフィトクロム遺伝子の探索を行い、1 分子種のフィトクロム
(MpPHY) をもつことを明らかにした。Mpphyの光に対する安定性および細胞内局在の解析
から、安定性は被子植物におけるⅡ型フィトクロムに、細胞内局在は被子植物におけるⅠ型
フィトクロムに類似した性質をもつことを明らかにした。野生型やMpPHY 過剰発現株では
無性芽の発芽や、LHCBおよび POR といった光誘導性遺伝子の発現が赤色光/遠赤色光可逆
的に制御されたのに対し、恒常的に活性型になると予想された点変異を導入したMpphyY241H
を発現する形質転換体ではそれらの応答の光依存性が喪失した。これらの結果から、Mpphy
はシロイヌナズナのフィトクロムと同様に赤色光依存的に遺伝子発現を制御することで生
理応答を制御することが示唆された。
次に、Mpphy を介した遺伝子発現制御に関与する候補遺伝子として PIF 相同遺伝子の探
索を行い、ゼニゴケが 1 分子種の PIF (MpPIF) をもつことを明らかにした。相同組み換え
による MpPIF ノックアウト株 (MppifKO) を用いた解析から、MppifKOでは MpphyY241H発現
株と同様に無性芽の発芽や光誘導性遺伝子の発現の光依存性が喪失することを見出し、
MpPIF が暗黒下で無性芽の発芽や光誘導性遺伝子の発現を抑制するフィトクロムシグナル
伝達の抑制因子であることを明らかにした。また生化学的な解析から、MpPIF が暗黒下で
蓄積し、赤色光依存的に分解されること、MpphyY241H発現株では暗黒下でも MpPIF が蓄積
しないこと、活性型Mpphyと APAモチーフを介して光依存的に相互作用することを明らか
にした。これらの結果から、MpPIFは赤色光依存的に活性型 Mpphyと相互作用することで
分解されることが示唆された。以上の結果から、基部陸上植物であるゼニゴケがフィトクロ
ムと PIF 転写因子との光依存的な相互作用および PIF 転写因子の分解による遺伝子発現制
御というシロイヌナズナと共通したフィトクロムシグナル伝達機構をもつことを明らかに
した。本研究の結果、PIFを介した転写制御による赤色光シグナル伝達機構は陸上植物進化
の基部に位置する苔類の時点で既に獲得された形質であることが示唆され、陸上植物の共
通祖先において獲得されていた可能性が考えられた。

3
略語表
PHY PHYTOCHROME
PIF PHYTOCHROME INTERACTING FACTOR
LHCB LIGHT-HARVESTING CHLOROPHYLL A/B-BINDING
POR PROTOCHLOROPHYLLIDE OXIDOREDUCTASE
FHY1 FAR-RED ELONGATED HYPOCOTYL1
FHL FHY1-LIKE
TOC1 TIMING OF CAB EXPRESSION 1
PIL PIF3-LIKE
COP1 CONSTITUTIVE PHOTOMORPHOGENIC 1
SPA SUPPRESSOR OF PHYA-105
HY5 ELONGATED HYPOCOTYL5
EF1 ELONGATION FACTOR 1
LRL1 LJRHL1-LIKE 1
APT ADENINE PHOSPHORIBOSYL TRANSFERASE
SPT SPATULA
ALC ALCATRAZ
UGPase UDP-glucose pyrophosphorylase
SCF Suppressor of kinetochore protein1, Cullin, Ring-Box1
CRISPR clustered regularly interspaced short palindromic repeats
CAS9 CRISPR associated proteins
R red light
FR far-red light
Pr red light absorbing
Pfr far-red light absorbing
DNA deoxyribonucleic acid
cDNA complementary DNA
RACE rapid amplification of cDNA ends
RNA ribonucleic acid
mRNA messenger RNA
PCR polymerase chain reaction
qRT-PCR quantitative reverse transcription PCR
APA active phytochrome A-binding
APB active phytochrome B-binding
PB phytochromobilin

4
VLFR very low fluence response
LFR low fluence response
HIR high irradiance response
bHLH basic helix-loop-helix
CaMV cauliflower mosaic virus
CBD chitin binding domain
EST expressed-sequence tag
EDTA ethylenediaminetetraacetic acid
DMSO dimethyl sulfoxide
BSA bovine serum albumin
PBS phosphate buffered saline
DTT dithiothreitol
IPTG isopropyl β-D-thiogalactopyranoside
PMSF phenylmethylsulfonyl fluoride
SDS sodium dodecyl sulfate
PAGE polyacrylamide gel electrophoresis
PVDF polyvinylidene difluoride
MUSCLE Multiple Sequence Comparison by Log-Expectation
JTT Jones-Thornton-Taylor
JGI Joint Genome Institute
BLAST Basic Local Alignment Search Tool
LED light emitting diode

5
序論
植物の環境応答
移動能をもたない植物にとって、周囲の環境に自身を適応させることは生存する上で必
要不可欠である。そのため、植物は光や温度、重力などを受容し、それらに従って最適な形
態形成や運動応答を行えるように進化を遂げてきた。様々な環境情報の中でも、「光」は光
合成のエネルギー源であると同時に、日長や周囲の植物体との位置関係を知るための環境
情報としても重要な因子であり、生活環を通して植物の生長、発生に多大な影響を与える因
子の 1つである。植物は光の強度、波長、方向、照射時間などを正確に感知し、自らの生体
情報に変換することで、種子の発芽、胚軸伸長阻害や子葉の展開といった光形態形成、成長
相転換といった組織レベルの形態形成から、光屈性や葉緑体定位運動といった細胞レベル
の運動応答まで、様々な生理応答を示すことが知られている。
光受容体フィトクロム
植物は光の情報を正確に感知するため、光の波長ごとに異なる光受容体をもっており、フ
ィトクロム、クリプトクロム、フォトトロピン、ZEITLUPE/FLAVIN-BINDING, KELCH
REPEAT, F-BOX1/LOV KELCH PROTEIN2、UV RESISTANCE LOCUS8がこれまでに同定さ
れている (Kami et al., 2010; Ito et al., 2012; Jenkins, 2014; Christie et al., 2015)。これらの光受容
体の中で、フィトクロムは赤色光および遠赤色光を受容する唯一の光受容体であり、その大
きな特徴として赤色光を受容することで生理学的に不活性型である Pr 型から活性型である
Pfr型へ、遠赤色光を受容することで Pfr型から Pr型へ光可逆的に構造変化する「光可逆性」
をもつ (図 1)。この性質によって、フィトクロムは光シグナルの ON/OFFを素早く切り替え
るスイッチとして機能することや、赤色光と遠赤色光の比を感知することが可能となって
いる (Mancinelli, 1994)。また、Pr型が Pfr型よりも熱力学的に安定なため、暗黒下では徐々
に Pfr型から Pr型へと変換される「暗反転」がみられる (Mancinelli, 1994)。
図 1 フィトクロムの光可逆的な構造変化の模式図

6
フィトクロムの分子構造は大きく N 末端領域と C 末端領域に分けることができる
(Nagatani, 2010)。N 末端領域には発色団結合ドメインである GAF (cGMP-specific
phosphodiesterases, adenylyl cyclases, FhlA) ドメインや、Pfr型の安定性に関与すると考えら
れる PHY (phytochrome-specific GAF-related ) ドメインなどが存在し、C末端領域には PAS
(PER, ARNT, SIM) ドメインやヒスチジンキナーゼ様ドメイン (histidine-kinase-related
domain; HKRD) が存在する。PASドメインや HKRD が存在することから、C 末端領域がシ
グナル伝達に重要であると長年考えられてきたが、N 末端領域にダイマー形成能と核移行
シグナルを付与した N末端 phytochrome B (phyB) が phyB 変異体を相補し、同様に N 末端
phytochrome A (phyA) が phyA変異体を部分的に相補したことから、N 末端領域がシグナル
伝達において重要であることが示されている (Matsushita et al., 2003; Oka et al., 2004; Mateos
et al., 2006)。このことから、C 末端領域には二量体化に必要なドメインや核移行シグナル
配列が存在すると考えられている (Muller et al., 2009)。また、N末端領域と C 末端領域の
結合が赤色光依存的に弱まることから、不活性型では N 末端領域が C 末端領域に結合して
核移行能を抑制しており、活性型になって構造が変化することで抑制が解除されて核内に
移行する可能性が考えられている (Chen et al., 2005)。
フィトクロム発色団と発色団生合成酵素遺伝子
フィトクロムアポタンパク質が光を吸収し得る機能的なタンパク質となるためには、発
色団と結合することが不可欠である。フィトクロムの発色団は開環状テトラピロール化合
物であり、陸上植物ではフィトクロモビリン (PB) を発色団として持っていることが知ら
れている。植物のフィトクロムホロタンパク質は GAFドメイン内に存在するシステイン残
基に発色団がチオエーテル結合によって共有結合している (Lagarias and Lagarias, 1989)。ま
た、発色団が結合するシステイン残基を欠失させても僅かに光受容能がみられることから、
非共有結合的に発色団が結合できる可能性も示唆されている (Oka et al., 2011)。PB の生合
成酵素遺伝子はシロイヌナズナ (Arabidopsis thaliana) において白色光下で胚軸が徒長する
hy1, hy2変異体から単離同定された (Muramoto et al., 1999; Kohchi et al., 2001)。
フィトクロムの機能による分類
被子植物のフィトクロムは暗所で高レベルに蓄積し、明所ですみやかに分解される光に
対して不安定なⅠ型フィトクロムと、明暗に大きく左右されずに存在する光に対して安定
なⅡ型フィトクロムに大別される。多くの植物種では、フィトクロムは遺伝子ファミリーを
形成しており、シロイヌナズナにおいては phyA~E の 5 分子種が同定されている (Clack et
al., 1994)。5分子種のフィトクロムのうち、phyA はⅠ型に、phyB~E はⅡ型フィトクロムに
分類される。この光に対する安定性に起因して、暗所では phyA が、明所では phyB が最も
多く蓄積することが示されている (Sharrock and Clack, 2002)。そのため、暗所における微弱
な光の受容には主に phyA が、明所における光の受容には主に phyB が機能していると考え

7
られている。イネ (Oryza sativa) やトウモロコシ (Zea mays) といった単子葉植物において
も光に対して不安定な phyA と光に対して安定な phyB の存在が報告されており、分解の速
度は植物種間でやや異なっているものの I 型フィトクロムは赤色光照射後 2~4 時間以内に
分解がみられる (Reddy and Sharma, 1998; Xie et al., 2014)。
シロイヌナズナのフィトクロムは光依存的に核内に移行して機能することが知られてい
る (Kircher et al., 1999; Yamaguchi et al., 1999; Huq et al., 2003; Klose et al., 2015)。この核移行
に関する分子機構もⅠ型とⅡ型のフィトクロムで異なっていることが明らかとなっている
(Nagatani, 2004; Fankhauser and Chen, 2008)。phyA の核移行は赤色光または遠赤色光のいず
れの照射によっても観察され、その核移行には FAR-RED ELONGATED HYPOCOTYL1
(FHY1)、FHY1-LIKE (FHL) といった因子を必要とすることが報告されている (Hiltbrunner
et al., 2006; Genoud et al., 2008; Rausenberger et al., 2011)。一方で、phyB は赤色光下でのみ効
率的に核内に移行することが報告されており (Yamaguchi et al., 1999; Oka et al., 2012)、核移
行に FHY1 や FHL を必要とせず、自身の核移行シグナルまたはフィトクロムシグナルに関
与する転写因子との結合を介して核内に移行しているのではないかと考えられている
(Chen et al., 2005; Pfeiffer et al., 2012)。核に移行する速度もⅠ型とⅡ型のフィトクロムで異な
っていることが知られており、phyA-GFP が光照射後 15 分以内と短時間で核移行が観察さ
れるのに対し (Kim et al., 2000)、phyB-GFP は赤色光照射後 2時間程度で核内に観察され始
めることが報告されている (Gil et al., 2000)。
フィトクロムに制御される生理応答
フィトクロムによって制御される生理応答は、その光量依存性や光可逆性、相反則の有無
によって低光量反応 (low fluence response; LFR)、超低光量反応 (very low fluence response;
VLFR)、高照射反応 (high irradiance response; HIR) の 3種類に分類される (Mancinelli, 1994;
Casal et al., 1998)。これらの応答の中で、LFR が最も広く知られており、典型的な赤色光/遠
赤色光可逆性や相反則を示す応答である。VLFR は波長の依存性がなく、非常に低い光量 (<
1 nmol m-2) のあらゆる波長の可視光で誘導され、光可逆性を示さないことから非常に少数
の Pfr 型のフィトクロムが応答を引き起こすのに十分である可能性が考えられている
(Shinomura et al., 1996)。HIR は長時間の連続した光照射が応答に必要であり、基本的には光
可逆性や相反則がみられないが、限られた厳密な実験条件下では相反則や光可逆性を示す
ことが報告されている (Shinomura et al., 2000)。シロイヌナズナではⅠ型フィトクロムであ
る phyA が VLFR と遠赤色光高照射反応 (FR-HIR) を制御し (Nagatani et al., 1993; Parks and
Quail, 1993; Whitelam et al., 1993; Shinomura et al., 1996)、Ⅱ型フィトクロムである phyB が
LFR と赤色光高照射反応 (R-HIR) を制御している (Mancinelli, 1994; Shinomura et al., 1996)。
フィトクロム相互作用因子とシグナル伝達機構
フィトクロムが受容した光情報を下流因子に伝達するには何らかのタンパク質との相互

8
作用が必要であることが考えられたため、これまでに多くの相互作用因子の探索が行われ
ている (Bae and Choi, 2008)。それらの相互作用因子の中には、phyA および phyB の両方に
結合する因子だけでなく、上述した FHY1 や FHL のように一方のみに結合する因子も存在
する。さらに、それらの中でも赤色光依存的に Pfr 型のフィトクロムに結合する因子と、
Pfr/Pr型の区別なく結合する因子が存在する。このようなフィトクロム相互作用因子の中で、
代表的な因子として PHYTOCHROME INTERACTING FACTOR (PIF) ファミリーが挙げら
れる (Leivar and Monte, 2014)。PIFファミリーは bHLH subfamily VII(a+b) に属する bHLH 型
の転写因子であり (Pires and Dolan, 2010)、phyB の C 末端領域を用いた Yeast two-hybrid 法
によって最初に PIF3 が単離された (Ni et al., 1998)。その後、phyB の N末端領域や全長を用
いて PIF3 との相互作用が見直され、PIF3 が赤色光依存的に Pfr 型の phyB に結合すること
や、その相互作用が光可逆的であることが明らかとなった (Ni et al., 1999)。さらに、赤色光
下で胚軸が短くなる srl2 (short under red-light 2) 変異体の原因遺伝子として PIF4 が単離さ
れ (Huq and Quail, 2002)、TIMING OF CAB EXPRESSION 1 (TOC1) と相互作用する因子の
中で PIF3と相同性の高い遺伝子として PIF3-LIKE 1 (PIL1)、PIL2、PIL5、PIL6が単離され
た (Yamashino et al., 2003)。その後、PIL遺伝子を含む bHLH サブファミリーと phyB の相互
作用解析が行われ、相互作用が確認された因子がそれぞれ PIF1/PIL5、PIF5/PIL6、PIF6/PIL2
と名付けられた (Huq et al., 2004; Khanna et al., 2004)。また、その後 bHLH72が PIF7 として
(Leivar et al., 2008)、bHLH16が PIF8 として新たに同定されている (Leivar and Quail, 2011)。
これらの PIFファミリーは N 末端側に APB (Active phytochrome B binding) モチーフと呼ば
れる領域が存在しており、その領域に点変異を入れると phyB との相互作用が阻害されるこ
とから、活性型フィトクロムとの相互作用部位であると考えられている (Khanna et al., 2004)。
さらに、イネにおける PIF ファミリーにも N 末端側に保存性の高い領域が存在するが、シ
ロイヌナズナの PIFファミリーのもつ APB モチーフとの相同性は低く、生物種間の APBモ
チーフの保存性は低いと考えられる (Nakamura et al., 2007)。また、PIF ファミリーの中で
PIF1、PIF3 の 2 分子種は Pfr 型の phyA とも相互作用することが示されており (Zhu et al.,
2000; Shimizu-Sato et al., 2002)、APA (Active phytochrome A binding) モチーフが存在すると考
えられているが、相互作用に必要な領域は未だ明確ではない (Al-Sady et al., 2006; Shen et al.,
2008)。PIF ファミリーは暗黒下や赤色光/遠赤色光比が低い環境下において多数の下流遺伝
子の発現制御を行うことで、フィトクロムシグナル伝達の負の制御因子として機能してい
る。光を受容して活性型となったフィトクロムは PIFと相互作用し、PIFのリン酸化とそれ
に続くユビキチン・プロテアソーム系による分解を引き起こす (図 2)。この光依存的な分解
による PIF蓄積量の変化が、種子の発芽、芽生えの脱黄化、胚軸伸長阻害、避陰応答といっ
た生理応答を引き起こすことが明らかとなっている (Leivar and Monte, 2014)。

9
図 2 PIFを介したフィトクロムシグナル伝達機構 (Castillon et al., 2007を参考に作製)
PIF以外の主要なフィトクロムシグナル伝達因子として、可視光のシグナル伝達において
中心的な抑制因子として機能する CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1) およ
び SUPPRESSOR OF PHYA-105 (SPA) ファミリーを介した経路が知られている (Huang et al.,
2014)。暗黒下において COP1/SPA複合体は ELONGATED HYPOCOTYL5 (HY5) のような光
形態形成の促進因子を分解することで生育に好ましくない環境下での光形態形成を抑制し
ている。近年の解析から、光を受容して活性型となったフィトクロムが SPA ファミリーと
光依存的に相互作用することで COP1/SPA 複合体の解離を促進することが報告されている
(Lu et al., 2015; Sheerin et al., 2015)。
上記のようにシロイヌナズナにおける知見は多く存在しているが、フィトクロム依存的
な遺伝子発現制御は様々な被子植物や (Sawers et al., 2002; Takano et al., 2005; Kebrom et al.,
2006; Takano et al., 2009; Kebrom et al., 2010)、裸子植物 (Alosi and Neale, 1992; Christensen et
al., 2002) においても報告されているのに対し、シロイヌナズナ以外の植物種におけるフィ
トクロムシグナルの分子機構に関する知見はほとんど存在しない。
基部植物のフィトクロムシグナル伝達機構
シダ植物やコケ植物蘚類といった基部植物においてもフィトクロムに関する知見が報告
されている (Suetsugu and Wada, 2007; Rosler et al., 2010)。シダ植物であるホウライシダ
(Adiantum capillus-veneris) では原糸体光屈性、葉緑体光定位運動など、被子植物では主にフ
ォトトロピンによって制御される応答が赤色光によって制御されることが報告されている
(Rosler et al., 2010; Jaedicke et al., 2012)。コケ植物では、蘚類に属するヒメツリガネゴケ
(Physcomitrella patens) やヤノウエノアカゴケ (Ceratodon purpureus) においてフィトクロム
に関する知見が報告されている。ヒメツリガネゴケには PpPHY1~4 および PpPHY5a~5c の
7 分子種が、ヤノウエノアカゴケには CpPHY1~4 の 4分子種が存在し、原糸体光屈性や分岐

10
の形成、葉緑体光定位運動を制御していることが報告されている (Pasentsis et al., 1998;
Mittmann et al., 2004; Uenaka et al., 2005; Mittmann et al., 2009)。これらのフィトクロムの中で、
Cpphy1 は C 末端に HKRD の代わりにセリン/スレオニンキナーゼと高い相同性を示す配列
をもった特殊な分子である (Thummler et al., 1992)。Cpphy1 以外は典型的な陸上植物フィト
クロムと共通したドメイン構造ではあるが、それらのエキソン・イントロン構造は被子植物
のフィトクロムと異なっていることが知られている (Mittmann et al., 2009)。また、Ppphy1~4
はプロトプラストを用いた一過的発現系において細胞質に局在することが示されており、
核移行シグナルを付加した Ppphy1 および Ppphy2 が葉緑体光定位運動に機能的でないこと
から、上記の応答は Ppphyの細胞質における機能であることが示唆されている (Uenaka and
Kadota, 2007)。このように基部植物のフィトクロムに関する知見は細胞質における機能が多
く存在するが、近年になって核における機能がいくつか報告されている。ホウライシダでは、
マイクロビームによって核内のフィトクロムを選択的に活性化することによって胞子発芽
を誘導できることが報告されている (Tsuboi et al., 2012)。ヒメツリガネゴケでは、連続遠赤
色光下において PpPHY1および PpPHY3が細胞質から核内へと移行し、FR-HIR 様の応答を
制御することが報告されている (Possart and Hiltbrunner, 2013)。しかしながら、基部植物に
おけるフィトクロムシグナル伝達の分子機構や遺伝子発現制御の有無は未だ直接的に示さ
れていない。近年の系統解析の結果から、被子植物や蘚類を始めとする多くの植物種では種
特異的な遺伝子重複によってフィトクロムの分子種が増加していることが示唆されている
(Li et al., 2015)。一方で、苔類やツノゴケ類では遺伝子重複がみられず、これらの種におけ
る 1 分子種のフィトクロムがもつ祖先的な性質を明らかにすることが、フィトクロム遺伝
子の分子進化を考えるうえで重要であると期待されている (Li et al., 2015)。
苔類ゼニゴケ (Marchantia polymorpha)
ゼニゴケは苔類に属するコケ植物で、その生活環の大部分を半数体である配偶体で過ご
すことが知られている (図 3)。葉状体 (Thallus) の背側には葉緑体に富んだ同化糸が存在す
る気室が存在し、無性生殖器官である杯状体 (Gemma cup) や無性芽 (Gemma) が形成され
る。葉状体の腹側には地表に密着するための仮根 (Rhizoid) や腹鱗片が形成される。ゼニゴ
ケは雌雄異株であり、造精器 (Antheridium) または造卵器 (Archegonium) をそれぞれの株が
形成することで有性生殖を行うことが知られている。ゼニゴケは陸上植物進化の基部に位
置すると考えられている苔類に属することから (Qiu et al., 2006)、植物の進化を考える上で
も非常に重要なモデルであり、被子植物に比べて体制が単純かつ遺伝子の冗長性が低いた
め、陸上植物の祖先的なシグナル伝達機構を備えている可能性が考えられる。従って、ゼニ
ゴケを用いた研究により、陸上植物に普遍的なシグナル伝達機構の根幹となる分子機構を
理解できると期待される。また、ゼニゴケは生活環の大半が半数体であるため、変異体・形
質転換体の表現型が当代で観察できる、無性生殖によってクローンを簡単に大量に増殖で
きるといった生物学的な利点を備えているだけでなく、Joint Genome Institute (JGI) による

11
ゲノム解読によりシグナル伝達に関与する遺伝子の冗長性が低いことが明らかとねってい
ること (未発表データ)、高効率の形質転換法や (Ishizaki et al., 2008)、相同組み換えによる
ジーンターゲティング(Ishizaki et al., 2013)、CRISPR/CAS9を用いたゲノム編集といった様々
な遺伝学的ツールが整備されており (Sugano et al., 2014)、基部陸上植物の新しいモデル植物
の 1つとなっている。
図 3 ゼニゴケの生活環

12
過去に、ゼニゴケの赤色光応答がいくつか報告されている。明暗周期で培養しているゼニ
ゴケの明期終了時に遠赤色光をパルス照射すると、葉状体の垂直方向への成長やクロロフ
ィル量の減少が観察され、これらの応答は遠赤色光の後に赤色光を照射することで打ち消
されたことからフィトクロムによる制御が示唆されている (Fredericq and De Greef, 1966;
Hartmann and Jenkins, 1984)。また、赤色光および遠赤色光のパルス照射を用いた解析から、
葉状体の老化や無性芽の発芽が赤色光/遠赤色光可逆性を示すことからフィトクロムによっ
て制御されることが示唆されている (Ninnemann and Halbsguth, 1965; De Greef et al., 1971;
Otto and Halbsguth, 1976)。さらに、ゼニゴケのフィトクロムが葉状体の切断片からの再生過
程において、細胞周期への再進入や再生芽の形態形成を制御することが近年報告されてい
る (Nishihama et al., 2015)。
本研究では、苔類ゼニゴケをモデルとしてフィトクロムの祖先的な性質を解析するとと
もに、そのシグナル伝達機構について解析することで、陸上植物のフィトクロムシグナル伝
達機構の進化を解明することを目的に研究を行い、ゼニゴケがそれぞれ 1 分子種のフィト
クロムと PIF転写因子をもつことを明らかにした。また、生化学的、細胞生物学的な解析か
ら、1分子種のフィトクロムが被子植物における I型およびⅡ型フィトクロムの特徴を併せ
もつことを明らかにした。さらに、陸上植物進化の基部に位置する苔類ゼニゴケが、フィト
クロムと PIF 転写因子の光依存的な相互作用および PIF の分解による遺伝子発現制御機構
をもつことを明らかとした。

13
結果
フィトクロム遺伝子の単離
ゼニゴケにおける赤色光シグナル伝達機構を明らかにするため、cDNAライブラリーおよ
びゲノムライブラリーを用いて赤色光受容体であるフィトクロム遺伝子の探索を行った。
その結果、ゼニゴケはゲノム上にフィトクロム相同遺伝子 (MpPHY) を 1 分子種のみもつ
ことが明らかとなった。得られたMpPHY のゲノム配列を調べたところ、陸上植物の典型的
なフィトクロム遺伝子とイントロン挿入位置が完全に保存されていた (図 4)。次に、アミノ
酸配列を比較したところ、陸上植物の典型的なフィトクロムと同じドメイン構造をもつこ
とが明らかとなった (図 4)。そこで、Mpphyの分光学的性質を明らかにするため、発色団と
して PB の付加した Mpphy の N 末端領域を大腸菌で発現、精製し、その吸収スペクトル
を測定した。その結果、組み換え Mpphyタンパク質は赤色光/遠赤色光可逆的な分光学的性
質を示した (図 5)。これらの結果から、Mpphy は陸上植物の典型的なフィトクロムと共通
した分光学的性質をもつ機能的なフィトクロムであることが示唆された。

14
図 4 ゼニゴケおよびシロイヌナズナのフィトクロムのマルチプルアライメント
赤い矢じりはイントロン挿入位置を示す。赤いアスタリスクは発色団結合残基を、青いアス
タリスクは光可逆的な構造変化に重要な残基を、マゼンタのアスタリスクは phyA における
ユビキチン結合残基を示す。赤、青、マゼンタ、黄、緑、紫の下線はそれぞれ N-terminal
extension,、PAS domain、GAF domain、PHY domain、PAS-related domain、histidine kinase-related
domain (HKRD) を示す。
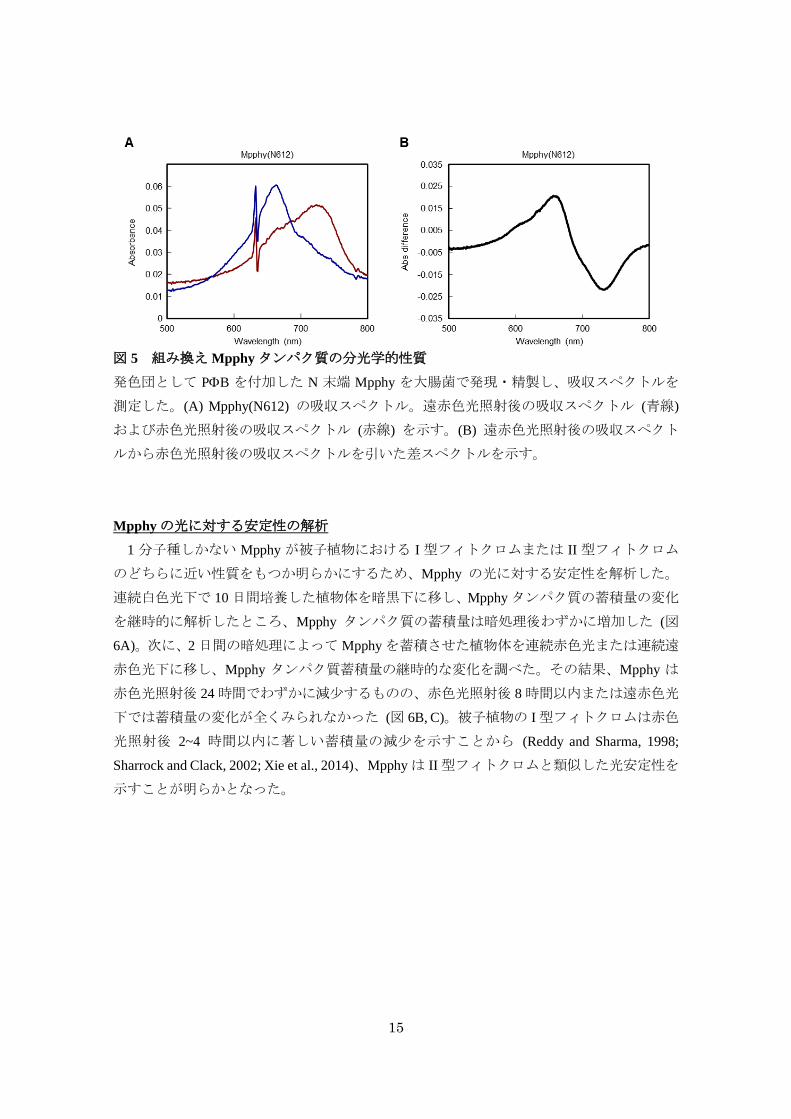
15
図 5 組み換え Mpphyタンパク質の分光学的性質
発色団として PB を付加した N 末端 Mpphy を大腸菌で発現・精製し、吸収スペクトルを
測定した。(A) Mpphy(N612) の吸収スペクトル。遠赤色光照射後の吸収スペクトル (青線)
および赤色光照射後の吸収スペクトル (赤線) を示す。(B) 遠赤色光照射後の吸収スペクト
ルから赤色光照射後の吸収スペクトルを引いた差スペクトルを示す。
Mpphyの光に対する安定性の解析
1 分子種しかない Mpphy が被子植物における I 型フィトクロムまたは II 型フィトクロム
のどちらに近い性質をもつか明らかにするため、Mpphy の光に対する安定性を解析した。
連続白色光下で 10日間培養した植物体を暗黒下に移し、Mpphyタンパク質の蓄積量の変化
を継時的に解析したところ、Mpphy タンパク質の蓄積量は暗処理後わずかに増加した (図
6A)。次に、2日間の暗処理によって Mpphyを蓄積させた植物体を連続赤色光または連続遠
赤色光下に移し、Mpphy タンパク質蓄積量の継時的な変化を調べた。その結果、Mpphy は
赤色光照射後 24時間でわずかに減少するものの、赤色光照射後 8時間以内または遠赤色光
下では蓄積量の変化が全くみられなかった (図 6B, C)。被子植物の I型フィトクロムは赤色
光照射後 2~4 時間以内に著しい蓄積量の減少を示すことから (Reddy and Sharma, 1998;
Sharrock and Clack, 2002; Xie et al., 2014)、Mpphyは II型フィトクロムと類似した光安定性を
示すことが明らかとなった。

16
図 6 Mpphyタンパク質の光に対する安定性の評価
抗 Mpphy 抗体を用いて野生型における内性 Mpphy タンパク質蓄積量の光に対する挙動を
解析した。連続白色光下で 10日間培養した植物体を次に示す光条件に移して解析を行った。
(A) 暗黒下、(B) 48時間の暗処理+連続赤色光 (30 µmol photons m-2 s-1)、(C) 48時間の暗処理
+連続遠赤色光 (30 µmol photons m-2 s-1)。抗 Histone H3抗体の結果をコントロールとして示
した。
Mpphyの細胞内局在解析
シロイヌナズナにおいて、I 型フィトクロムである phyA は赤色光および遠赤色光のいず
れの照射においても細胞質から核内への移行がみられるのに対し、II型フィトクロムである
phyB は赤色光照射下でのみ効率的な核移行がみられることが知られている (Kircher et al.,
1999; Yamaguchi et al., 1999)。そこで、Mpphyの細胞内局在パターンが I型および II型のど
ちらに近いのか明らかにするため、Mpphyの C 末端に蛍光タンパク質 Citrine を付加した融
合タンパク質を構成的発現プロモーターである CaMV 35S プロモーター (pro35S) の制御下
で発現するコンストラクトを導入した形質転換体 (pro35S:MpPHY-Citrine) を作出した。
pro35S:MpPHY-Citrine 株の無性芽を連続白色光下で 1 日間培養後、6 日間の暗処理を施し、
暗処理直後または暗処理後に赤色光または遠赤色光を 10 分間照射して蛍光観察を行った。
その結果、Mpphy-Citrine の蛍光は暗黒下では細胞の周縁部でのみ観察されたのに対し (図
7)、赤色光または遠赤色光照射後には主に核での蛍光が観察された (図 7)。

17
図 7 Mpphy-Citrineの細胞内局在解析
pro35S:MpPHY-Citrine株を連続白色光下で 1日間培養後、暗黒下に 6日間移したサンプル (上
段) および暗処理後に連続赤色光 (30 µmol photons m-2 s-1; 中段) または連続遠赤色光 (30
µmol photons m-2 s-1; 下段) を 10分間照射したサンプルの蛍光観察。スケールバーは 20 µm。
また、光可逆的な構造変化に重要なチロシン残基 (Tyr241) をヒスチジン残基に置換し、
恒常的に活性型として機能することが予想される 1アミノ酸置換体 (MpphyY241H) を用いて
同様の解析を行ったところ、MpphyY241H-Citrine は暗黒下においても核での蛍光が観察され
た (図 8)。組み換えMpphyY241Hタンパク質は phyBY276Hと同様に光可逆的な吸光スペクトル
の変化が完全に喪失したことから (図 9)、phyBY276H と同様に生体内で活性型として機能す
る可能性が考えられた (Su and Lagarias, 2007)。これらの結果から、Mpphyは光照射によっ
て活性型になると核内へと移行することが示唆された。
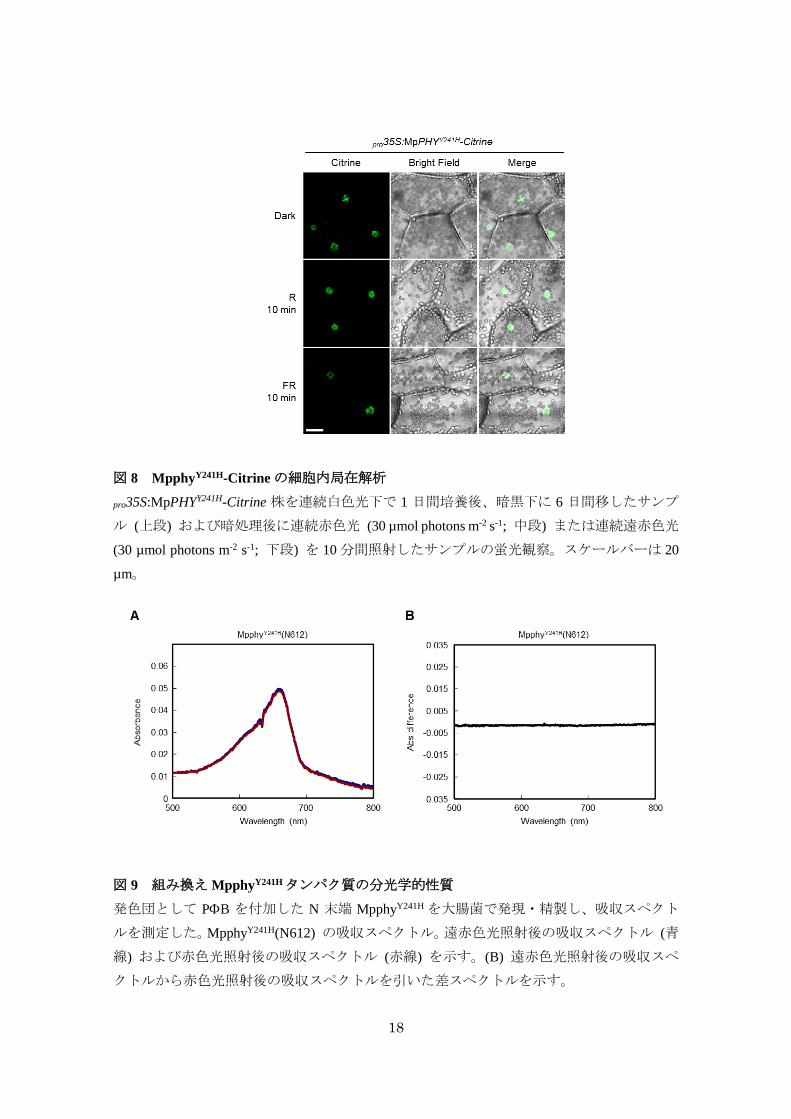
18
図 8 MpphyY241H-Citrineの細胞内局在解析
pro35S:MpPHYY241H-Citrine 株を連続白色光下で 1 日間培養後、暗黒下に 6 日間移したサンプ
ル (上段) および暗処理後に連続赤色光 (30 µmol photons m-2 s-1; 中段) または連続遠赤色光
(30 µmol photons m-2 s-1; 下段) を 10分間照射したサンプルの蛍光観察。スケールバーは 20
µm。
図 9 組み換え MpphyY241Hタンパク質の分光学的性質
発色団として PB を付加した N 末端 MpphyY241H を大腸菌で発現・精製し、吸収スペクト
ルを測定した。MpphyY241H(N612) の吸収スペクトル。遠赤色光照射後の吸収スペクトル (青
線) および赤色光照射後の吸収スペクトル (赤線) を示す。(B) 遠赤色光照射後の吸収スペ
クトルから赤色光照射後の吸収スペクトルを引いた差スペクトルを示す。

19
光依存的な Mpphy の細胞内局在変化をさらに確認するため、細胞分画による生化学的手
法によって内性Mpphyタンパク質の細胞内局在を解析した。連続白色光下で 10日間培養し
た植物体に対して 2日間の暗処理を施し、赤色光または遠赤色光を照射して 10分および 60
分後のサンプルについて細胞分画を行い、ウエスタンブロットによって細胞内局在を解析
した。その結果、蛍光観察の結果と一致して赤色光または遠赤色光照射のいずれにおいても
10 分後に核内での Mpphy 蓄積量が増加した (図 10)。しかし、光照射に対する迅速な核移
行は蛍光観察と一致していたが、細胞分画の結果から大部分の Mpphy タンパク質は光照射
後も細胞質に残っていることが明らかとなった (図 10)。さらに、遠赤色光照射サンプルの
核画分において移動度が変化したバンドが確認されたことから (図 10B)、遠赤色光照射下
での核内のフィトクロムは何らかの翻訳後修飾を受けることが明らかとなった。以上の結
果から、Mpphy は赤色光および遠赤色光のいずれの照射でも核内に移行するという被子植
物の I型フィトクロムと同様の細胞内局在パターンを示すことが明らかとなった。
図 10 細胞分画による内性 Mpphyタンパク質の細胞内局在解析
細胞分画およびウエスタンブロットによる細胞内局在解析。連続白色光下で 10日間培養し
た植物体に 48時間の暗処理を施した後、連続赤色光 (30 µmol photons m-2 s-1; A) または連
続遠赤色光 (30 µmol photons m-2 s-1; B) 照射下に移し、図に示した時間でサンプリングを行
い、細胞質画分 (Nucleus-depleted) および核画分 (Nucleus-enriched) を用いてウエスタンブ
ロットを行った。抗 UGPase 抗体を細胞質マーカーとして、抗 Histone H3 抗体を核マーカ
ーとして用いた。

20
Mpphyが制御する生理応答の解析
過去に、ゼニゴケの無性芽が赤色光依存的に発芽し、遠赤色光によって発芽が抑制される
ことから、無性芽の発芽がフィトクロムによって制御されることが示唆されている (Otto
and Halbsguth, 1976)。そこで、野生型の MpPHY 遺伝子を構成的発現プロモーターである
ELONGATION FACTOR 1 (MpEF1) プロモーターの制御下で発現するコンストラクトを導
入した MpPHY 過剰発現株 (proMpEF1:MpPHY) および恒常的に活性型として機能すること
が予想される点変異をもつ MpPHYY241H を MpEF1 プロモーターの制御下で発現するコンス
トラクトを導入した恒常活性型 MpPHY 発現株 (proMpEF1:MpPHYY241H) を用いて、Mpphyが
無性芽の発芽に関与するかどうかを解析した。緑色安全光下で無性芽を寒天培地に播き、暗
黒下で 1日間吸水させた後、様々な光処理を施して発芽した個体数をカウントした。その結
果、野生型の無性芽では過去の報告と一致して赤色光のパルス照射によって発芽が誘導さ
れ、赤色光に続く遠赤色光によって発芽が阻害される赤色光/遠赤色光可逆性が見られた
(図 11A)。また、赤色光によって発芽が誘導された個体数は光量依存的に増加した (図 11B)。
さらに、遠赤色光のパルス照射や連続遠赤色光は無性芽の発芽に有効でないことが明らか
となった (図 11A)。これらの結果から、ゼニゴケの無性芽の発芽は VLFR や FR-HIR ではな
く、典型的な光可逆性を示す LFR に分類されると考えられた。proMpEF1:MpPHY では赤色
光によって発芽誘導される個体数が野生型と比較して僅かに増加したが、遠赤色光による
発芽の阻害は野生型と同程度にみられた (図 11A)。対照的に、proMpEF1:MpPHYY241H では、
ほぼ全ての個体が暗黒下においても発芽し、遠赤色光による発芽の阻害が全くみられなか
った (図 11A)。qRT-PCR によって proMpEF1:MpPHYY241Hおよび proMpEF1:MpPHY における
MpPHY 発現量を調べたところ、各形質転換体における MpPHY 発現量にほとんど差がみら
れなかったことから (図 12)、proMpEF1:MpPHYY241Hの表現型は発現量の差ではなく、アミノ
酸置換の影響であることが示された。これらの結果から、過去の観察結果と一致して
(Nishihama et al., 2015)、MpphyY241Hが生体内で恒常的に活性型として機能することが強く支
持された。
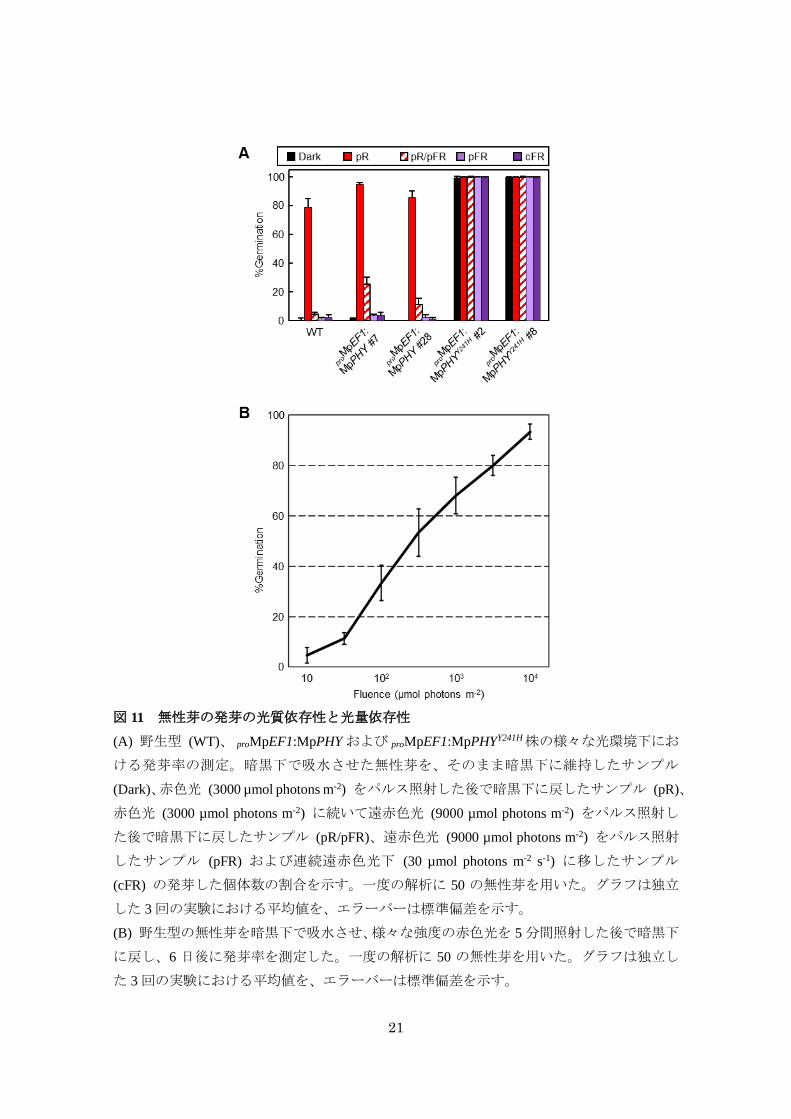
21
図 11 無性芽の発芽の光質依存性と光量依存性
(A) 野生型 (WT)、 proMpEF1:MpPHY および proMpEF1:MpPHYY241H株の様々な光環境下にお
ける発芽率の測定。暗黒下で吸水させた無性芽を、そのまま暗黒下に維持したサンプル
(Dark)、赤色光 (3000 µmol photons m-2) をパルス照射した後で暗黒下に戻したサンプル (pR)、
赤色光 (3000 µmol photons m-2) に続いて遠赤色光 (9000 µmol photons m-2) をパルス照射し
た後で暗黒下に戻したサンプル (pR/pFR)、遠赤色光 (9000 µmol photons m-2) をパルス照射
したサンプル (pFR) および連続遠赤色光下 (30 µmol photons m-2 s-1) に移したサンプル
(cFR) の発芽した個体数の割合を示す。一度の解析に 50 の無性芽を用いた。グラフは独立
した 3回の実験における平均値を、エラーバーは標準偏差を示す。
(B) 野生型の無性芽を暗黒下で吸水させ、様々な強度の赤色光を 5分間照射した後で暗黒下
に戻し、6 日後に発芽率を測定した。一度の解析に 50 の無性芽を用いた。グラフは独立し
た 3回の実験における平均値を、エラーバーは標準偏差を示す。
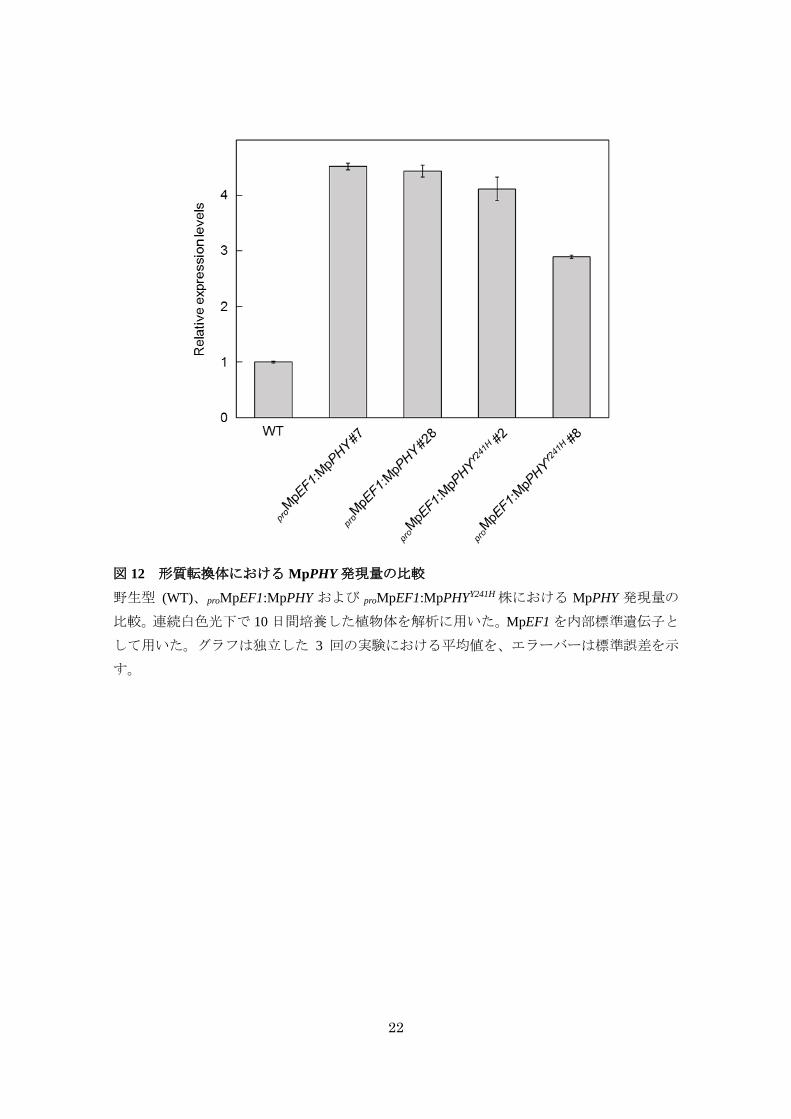
22
図 12 形質転換体における MpPHY発現量の比較
野生型 (WT)、proMpEF1:MpPHY および proMpEF1:MpPHYY241H株における MpPHY 発現量の
比較。連続白色光下で 10日間培養した植物体を解析に用いた。MpEF1を内部標準遺伝子と
して用いた。グラフは独立した 3 回の実験における平均値を、エラーバーは標準誤差を示
す。
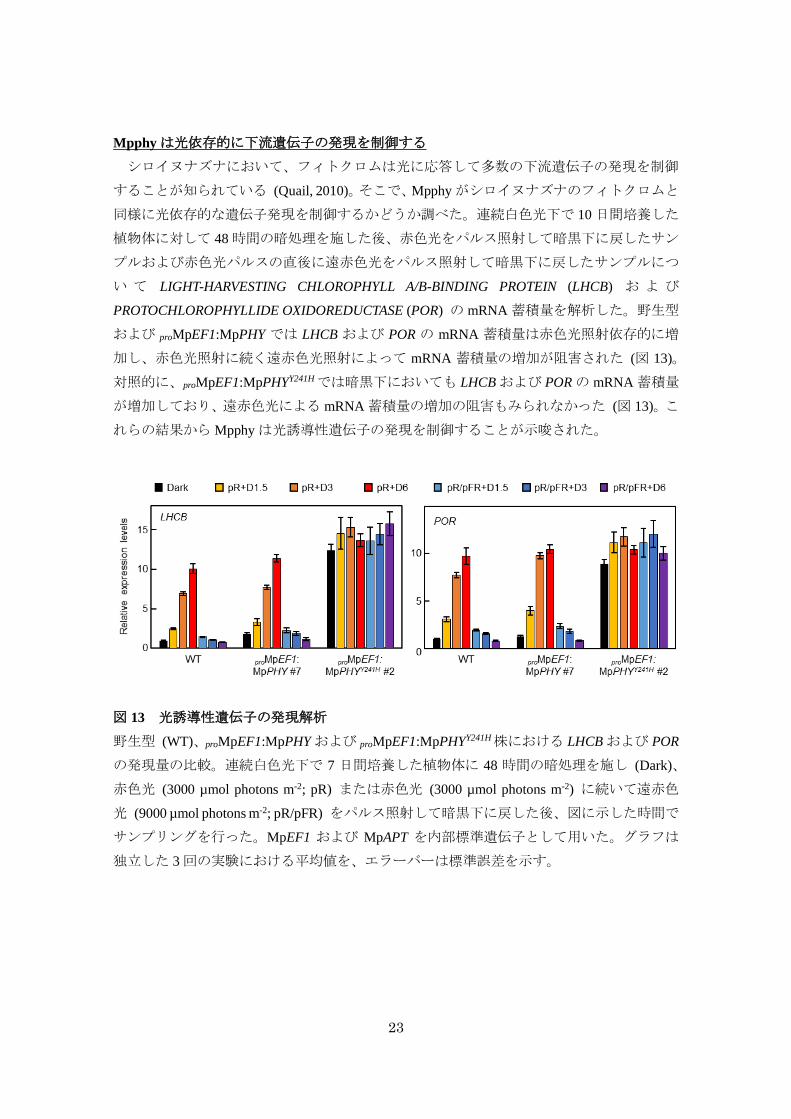
23
Mpphyは光依存的に下流遺伝子の発現を制御する
シロイヌナズナにおいて、フィトクロムは光に応答して多数の下流遺伝子の発現を制御
することが知られている (Quail, 2010)。そこで、Mpphyがシロイヌナズナのフィトクロムと
同様に光依存的な遺伝子発現を制御するかどうか調べた。連続白色光下で 10日間培養した
植物体に対して 48時間の暗処理を施した後、赤色光をパルス照射して暗黒下に戻したサン
プルおよび赤色光パルスの直後に遠赤色光をパルス照射して暗黒下に戻したサンプルにつ
い て LIGHT-HARVESTING CHLOROPHYLL A/B-BINDING PROTEIN (LHCB) お よ び
PROTOCHLOROPHYLLIDE OXIDOREDUCTASE (POR) の mRNA 蓄積量を解析した。野生型
および proMpEF1:MpPHY では LHCB および POR の mRNA 蓄積量は赤色光照射依存的に増
加し、赤色光照射に続く遠赤色光照射によって mRNA 蓄積量の増加が阻害された (図 13)。
対照的に、proMpEF1:MpPHYY241Hでは暗黒下においても LHCBおよび PORの mRNA蓄積量
が増加しており、遠赤色光による mRNA蓄積量の増加の阻害もみられなかった (図 13)。こ
れらの結果からMpphyは光誘導性遺伝子の発現を制御することが示唆された。
図 13 光誘導性遺伝子の発現解析
野生型 (WT)、proMpEF1:MpPHY および proMpEF1:MpPHYY241H株における LHCBおよび POR
の発現量の比較。連続白色光下で 7 日間培養した植物体に 48 時間の暗処理を施し (Dark)、
赤色光 (3000 µmol photons m-2; pR) または赤色光 (3000 µmol photons m-2) に続いて遠赤色
光 (9000 µmol photons m-2; pR/pFR) をパルス照射して暗黒下に戻した後、図に示した時間で
サンプリングを行った。MpEF1 および MpAPT を内部標準遺伝子として用いた。グラフは
独立した 3回の実験における平均値を、エラーバーは標準誤差を示す。

24
ゼニゴケは PIF転写因子を 1分子種のみもつ
これまでの解析から、フィトクロム依存的な遺伝子発現制御がゼニゴケにおいてもみら
れたため、次にMpphyシグナルに関与する転写因子を探索した。PIFファミリーは bHLH 転
写因子の Subfamily VII(a+b) に属することが報告されている (Pires and Dolan, 2010)。ゼニゴ
ケゲノムデータベースに対する BLAST および系統解析の結果から、ゼニゴケはシロイヌナ
ズナやイネの PIF ファミリーと同じクレードに属する遺伝子を 1 分子種のみもつことが明
らかとなった (図 14)。この候補遺伝子 (MpPIF) は C 末端領域に bHLH ドメインが非常に
高度に保存されており、シロイヌナズナの PIF1 および PIF3 に存在する phyA との相互作用
に重要であると予想される APAモチーフをもつことが明らかとなった (図 15)。しかしなが
ら、MpPIFはシロイヌナズナの全ての PIFファミリー転写因子に存在する phyB との相互作
用に重要な APBモチーフをもたないことが明らかとなった (図 15)。また、ヒメツリガネゴ
ケにおける PIF 相同遺伝子も APB モチーフをもたず、APA モチーフのみをもつことから
(図 16)、APB モチーフは陸上植物進化の過程で獲得された形質であることが示唆された。
また、BLAST 解析の結果、MpPIFと最も類似性の高かった Mapoly0502s0001 は、根毛形成
に関与することが報告されている LRL1 と同じクレード (Subfamily XI) に属した (Karas et
al., 2009)。Subfamily XIに属する遺伝子には APAモチーフや APB モチーフがみつからなか
ったことから、ゼニゴケが PIF転写因子を 1分子種のみもつことが強く支持された。
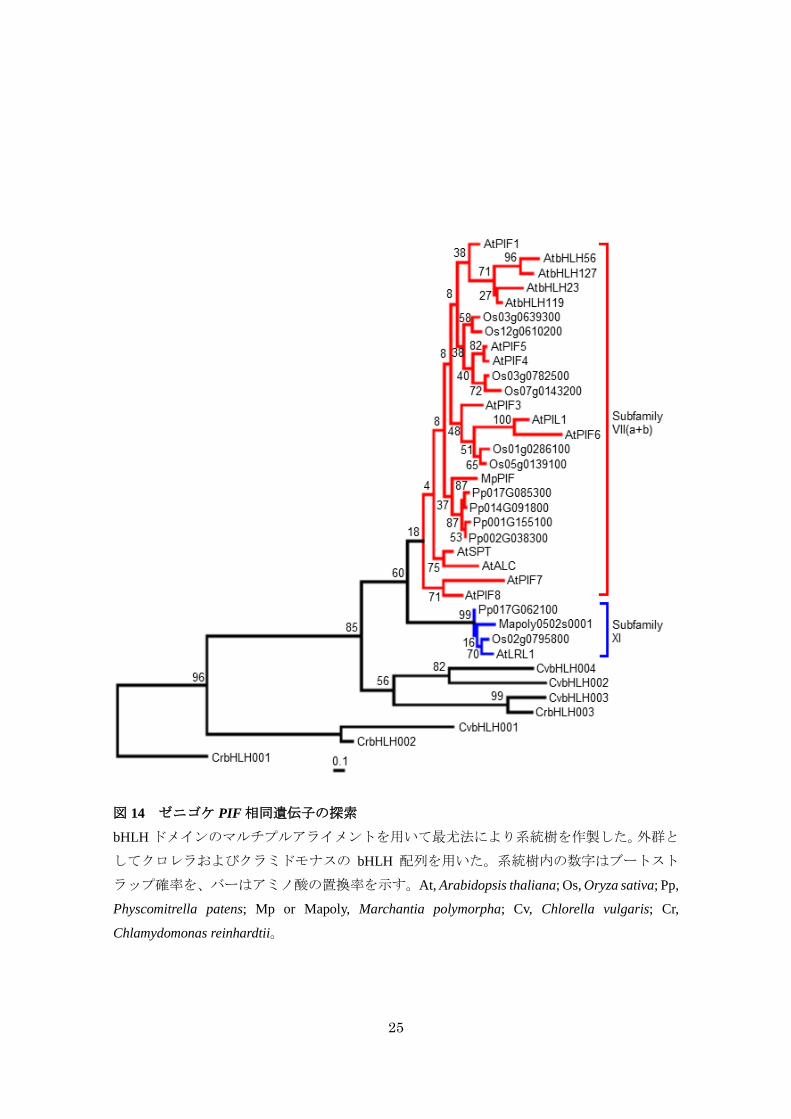
25
図 14 ゼニゴケ PIF相同遺伝子の探索
bHLH ドメインのマルチプルアライメントを用いて最尤法により系統樹を作製した。外群と
してクロレラおよびクラミドモナスの bHLH 配列を用いた。系統樹内の数字はブートスト
ラップ確率を、バーはアミノ酸の置換率を示す。At, Arabidopsis thaliana; Os, Oryza sativa; Pp,
Physcomitrella patens; Mp or Mapoly, Marchantia polymorpha; Cv, Chlorella vulgaris; Cr,
Chlamydomonas reinhardtii。

26
図 15 MpPIFとシロイヌナズナの PIFファミリーのマルチプルアライメント
赤とマゼンタの下線はそれぞれ APBモチーフと bHLH ドメインを示す。青枠は推定 APAモ
チーフを示す。赤いアスタリスクは PIFファミリーと phyB の結合に重要な残基を示す。青
のアスタリスクは phyA と PIF1 の結合に重要な残基を、マゼンタのアスタリスクは phyA と
PIF3 の結合に重要な残基を示す。

27
図 16 APAモチーフおよび APBモチーフのマルチプルアライメント
MpPIFとシロイヌナズナ、イネ、ヒメツリガネゴケの PIF相同遺伝子における APAモチー
フ (A) および APBモチーフ (B) のアミノ酸配列の比較。青のアスタリスクは phyAと PIF1
の結合に重要な残基を、マゼンタのアスタリスクは phyA と PIF3 の結合に重要な残基を示
す。黒のアスタリスクは高度に保存された残基を示す。赤のアスタリスクは PIFファミリー
と phyB の結合に重要な残基を示す。
MpPIFは Mpphyシグナル伝達の抑制因子である
MpPIFが Mpphyシグナルに関与するかどうかを明らかにするため、相同組み換えによる
MpPIF ノックアウト株 (MppifKO) を作出し (図 17)、前述した無性芽の発芽の表現型につい
て解析した。その結果、MppifKOでは、proMpEF1:MpPHYY241Hと同様に光条件に関わらずほぼ
全ての無性芽が発芽した (図 18A)。さらに、この発芽の光依存性の喪失は、MpPIF ゲノム
断片を導入した相補株 (proMpPIF:MpPIF/MppifKO) で野生型と同程度に回復したことから
(図 18A)、MpPIF がMppifKOにおける発芽の光依存性の喪失の責任遺伝子であることが示さ
れた。
次に、Mpphy 依存的な LHCB および POR の発現制御における MpPIF の機能について解
析した。その結果、MppifKOでは赤色光または遠赤色光パルス照射後に徐々に発現量が減少
するものの、暗黒下においても LHCB および POR の発現量が野生型より増加していた (図
18B)。また、相補株では野生型と同程度まで LHCB および PORの発現パターンが回復した
(図 18B)。以上の結果から、MpPIF は Mpphy シグナル伝達経路において無性芽の発芽およ
び光誘導性遺伝子の発現を負に制御することが示唆された。
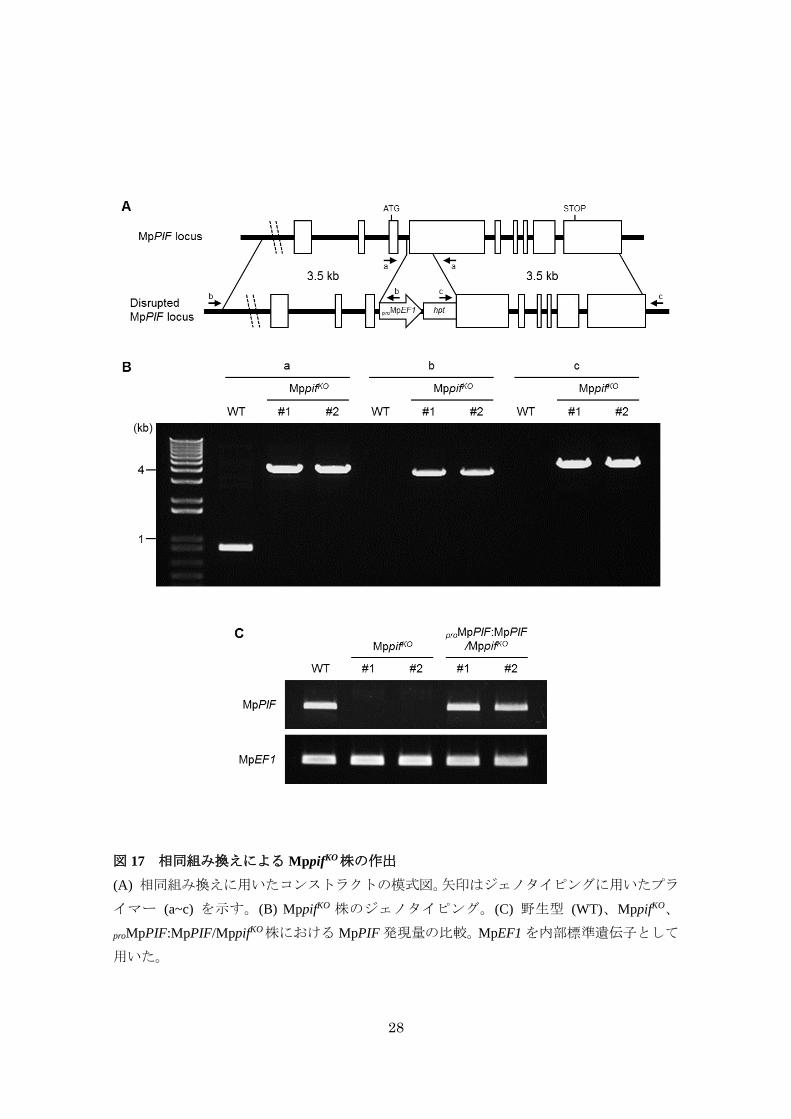
28
図 17 相同組み換えによる MppifKO株の作出
(A) 相同組み換えに用いたコンストラクトの模式図。矢印はジェノタイピングに用いたプラ
イマー (a~c) を示す。(B) MppifKO 株のジェノタイピング。(C) 野生型 (WT)、MppifKO、
proMpPIF:MpPIF/MppifKO株におけるMpPIF発現量の比較。MpEF1を内部標準遺伝子として
用いた。
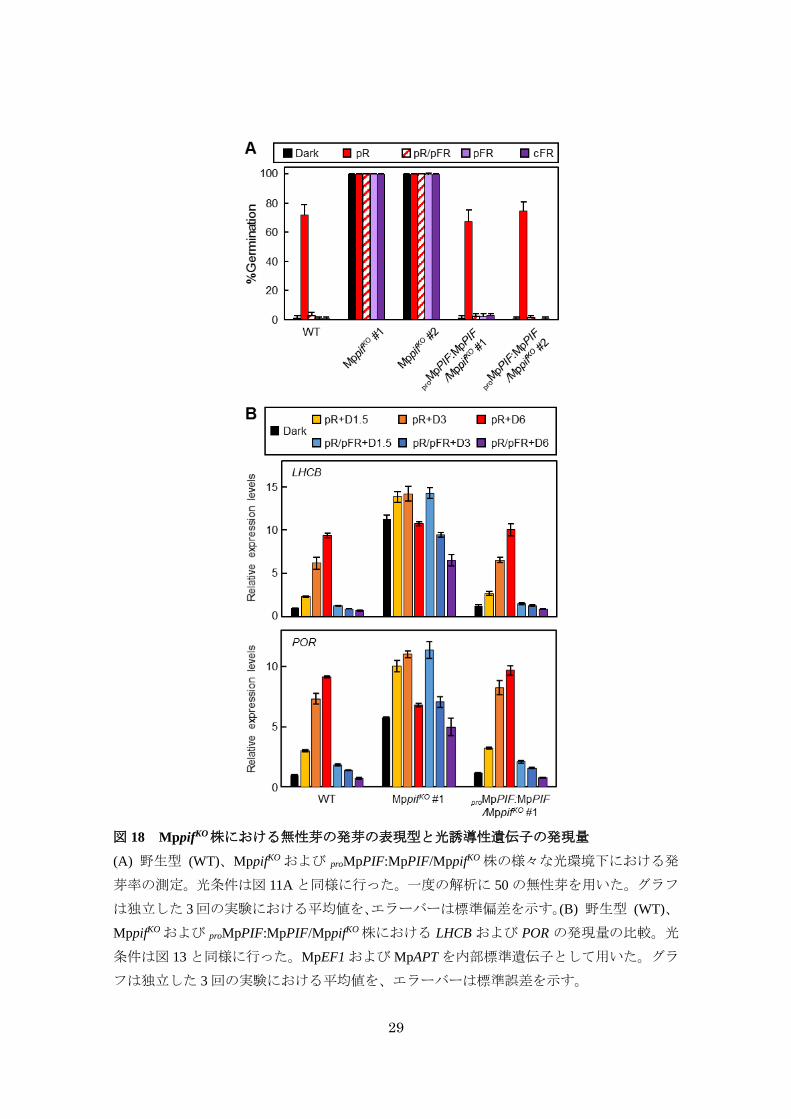
29
図 18 MppifKO株における無性芽の発芽の表現型と光誘導性遺伝子の発現量
(A) 野生型 (WT)、MppifKOおよび proMpPIF:MpPIF/MppifKO株の様々な光環境下における発
芽率の測定。光条件は図 11Aと同様に行った。一度の解析に 50の無性芽を用いた。グラフ
は独立した 3回の実験における平均値を、エラーバーは標準偏差を示す。(B) 野生型 (WT)、
MppifKOおよび proMpPIF:MpPIF/MppifKO株における LHCB および POR の発現量の比較。光
条件は図 13と同様に行った。MpEF1およびMpAPT を内部標準遺伝子として用いた。グラ
フは独立した 3回の実験における平均値を、エラーバーは標準誤差を示す。

30
Mpphyは光依存的に MpPIFタンパク質蓄積量を制御する
MpPIFが Mpphyシグナル伝達経路における遺伝子発現制御に関与することが示唆された
ため、光依存的な遺伝子発現制御の分子機構について解析した。シロイヌナズナにおいて、
活性型のフィトクロムが PIF ファミリー転写因子と相互作用することで、PIF のユビキチ
ン・プロテアソーム系による分解を誘起することが知られている (Al-Sady et al., 2006; Shen
et al., 2007; Shen et al., 2008)。そこで、まずMpPIFタンパク質蓄積量の光に対する挙動につ
いて解析した。連続白色光下で 10 日間培養した植物体に対して 48 時間の暗処理を施した
後、連続赤色光あるいは連続遠赤色光下に移して MpPIF タンパク質蓄積量について解析し
た。その結果、MpPIF タンパク質は連続白色光下の植物体ではほとんど検出されなかった
のに対し、暗処理後 24時間以降で顕著に蓄積量が増加した (図 19A)。また、この蓄積量の
増加は、暗処理直前の遠赤色光照射によって促進された (図 19B)。これは遠赤色光によっ
て Mpphy が速やかに不活性型へと変化したことによるものと考えられ、活性型 Mpphy が
MpPIF タンパク質の蓄積を阻害する可能性が考えられた。暗黒下で蓄積した MpPIF タンパ
ク質は赤色光照射によって 30分以内に速やかに分解されたのに対して (図 19C)、遠赤色光
照射ではタンパク質蓄積量に変化がみられなかった (図 19D)。赤色光照射条件において
MpPIF の mRNA 発現量には変動がみられなかったことから (図 20)、MpPIF タンパク質蓄
積量の速やかな減少はタンパク質レベルでの制御であることが示唆された。さらに、26Sプ
ロテアソームの阻害剤である MG132 の処理によって、赤色光による MpPIF の蓄積量の減
少が阻害されたことから (図 19E)、赤色光下での MpPIF 蓄積量の減少にユビキチン・プロ
テアソーム系が関与することが示唆された。また、proMpEF1:MpPHYY241Hでは暗黒下におい
てもMpPIFタンパク質の蓄積がみられなかった (図 21)。以上の結果から、赤色光下におけ
るMpPIFタンパク質の速やかな分解は活性型Mpphyによって引き起こされることが示唆さ
れた。
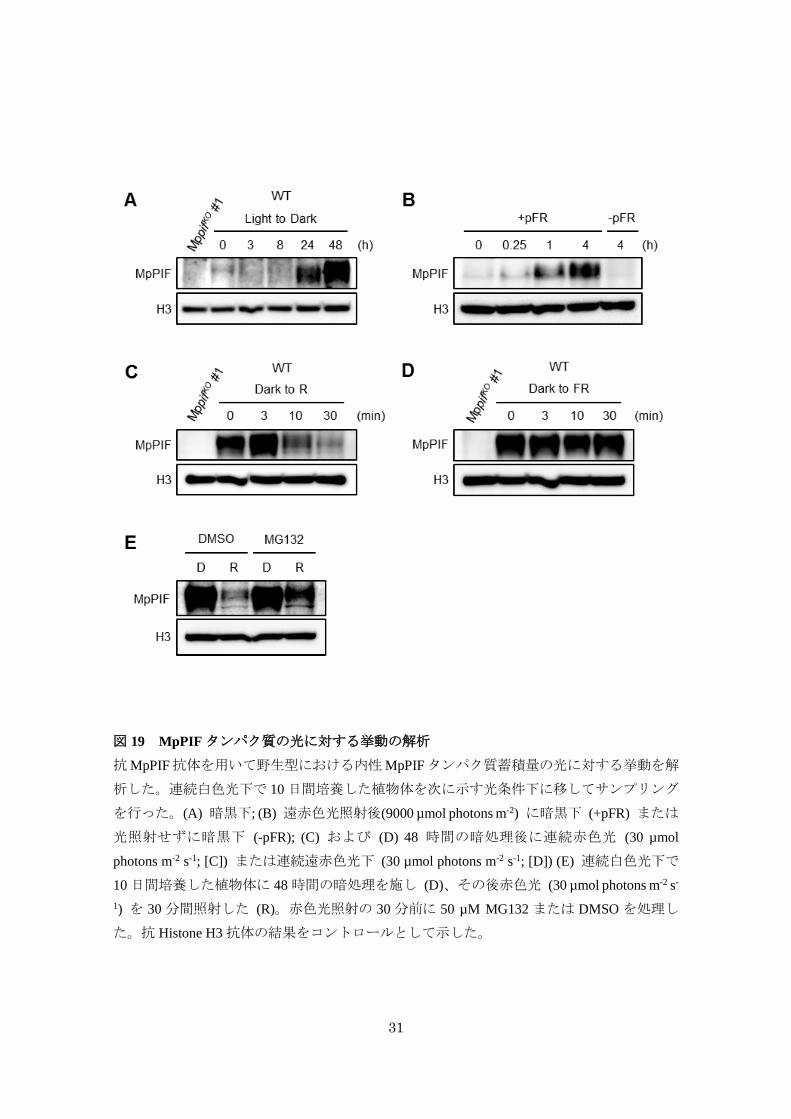
31
図 19 MpPIFタンパク質の光に対する挙動の解析
抗MpPIF抗体を用いて野生型における内性MpPIFタンパク質蓄積量の光に対する挙動を解
析した。連続白色光下で 10日間培養した植物体を次に示す光条件下に移してサンプリング
を行った。(A) 暗黒下; (B) 遠赤色光照射後(9000 µmol photons m-2) に暗黒下 (+pFR) または
光照射せずに暗黒下 (-pFR); (C) および (D) 48 時間の暗処理後に連続赤色光 (30 µmol
photons m-2 s-1; [C]) または連続遠赤色光下 (30 µmol photons m-2 s-1; [D]) (E) 連続白色光下で
10 日間培養した植物体に 48時間の暗処理を施し (D)、その後赤色光 (30 µmol photons m-2 s-
1) を 30 分間照射した (R)。赤色光照射の 30 分前に 50 µM MG132 または DMSO を処理し
た。抗 Histone H3 抗体の結果をコントロールとして示した。
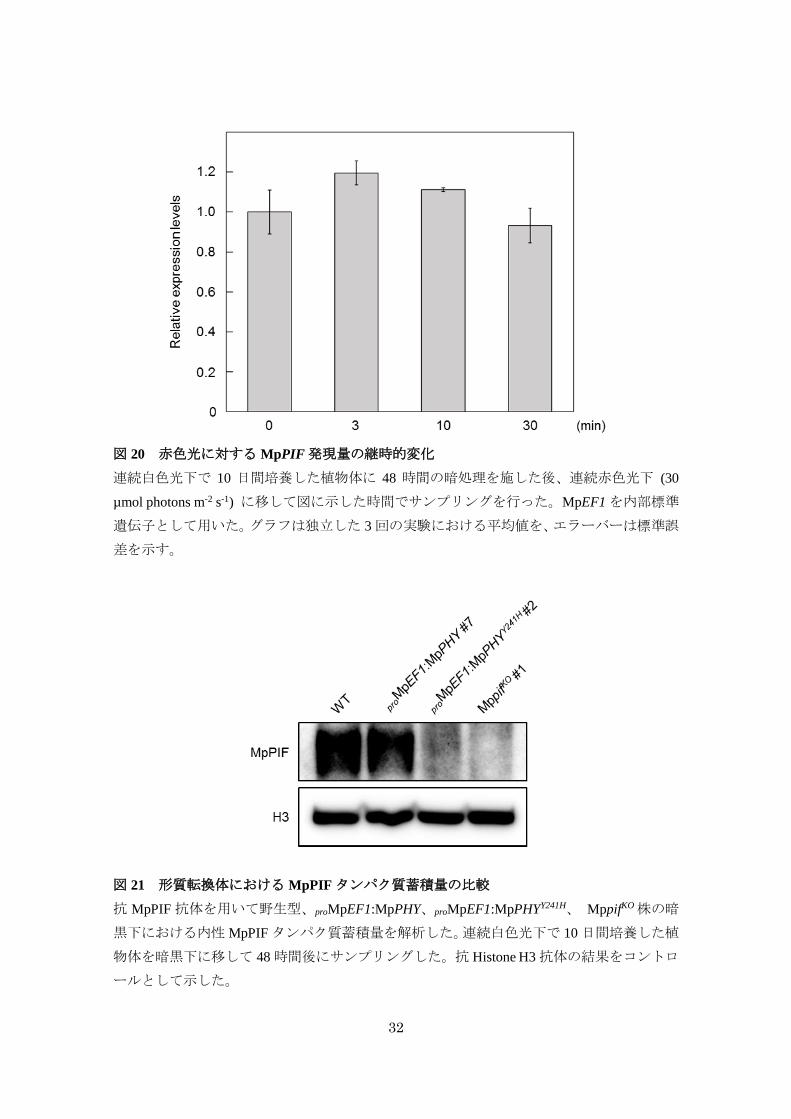
32
図 20 赤色光に対する MpPIF発現量の継時的変化
連続白色光下で 10 日間培養した植物体に 48 時間の暗処理を施した後、連続赤色光下 (30
µmol photons m-2 s-1) に移して図に示した時間でサンプリングを行った。MpEF1を内部標準
遺伝子として用いた。グラフは独立した 3回の実験における平均値を、エラーバーは標準誤
差を示す。
図 21 形質転換体における MpPIFタンパク質蓄積量の比較
抗 MpPIF 抗体を用いて野生型、proMpEF1:MpPHY、proMpEF1:MpPHYY241H、 MppifKO株の暗
黒下における内性MpPIFタンパク質蓄積量を解析した。連続白色光下で 10日間培養した植
物体を暗黒下に移して 48時間後にサンプリングした。抗 Histone H3抗体の結果をコントロ
ールとして示した。

33
続いて、Mpphyと MpPIF の相互作用および、その光依存性について解析した。大腸菌で
発現・精製した N 末端 Mpphy-PB と無細胞タンパク質発現系で調整した FLAG-MpPIF を
用いて in vitro プルダウン解析を行った。その結果、活性型の Mpphy は FLAG-MpPIF と相
互作用を示したのに対し、不活性型の Mpphyや MpPHYアポタンパク質は FLAG-MpPIFと
相互作用を示さなかった (図 22A)。次に、活性型 Mpphy と MpPIF の相互作用に APA モチ
ーフが必要であるかどうか調べるため、APA モチーフ内の 2 つのフェニルアラニン残基を
アラニンに置換した MpPIFF297A, F300A を作出し、Mpphy との相互作用を解析したところ、
MpPIFF297A, F300Aは活性型Mpphyと相互作用を示さなかった (図 22B)。このことから、APA
モチーフは Mpphy との相互作用に必要であることが示唆された。次に、生体内における
MpphyとMpPIFの光依存的な相互作用を明らかにするため、3FLAGタグを付加したMpPIF
を MppifKO に導入した proMpPIF:MpPIF-3FLAG/MppifKO を用いて共免疫沈降による相互作
用解析を行った。連続白色光下で 10日間培養した植物体に対して 48時間の暗処理を施し、
赤色光のみを照射した植物体および赤色光照射後に遠赤色光を照射した植物体を用いて、
FLAG 抗体による共免疫沈降を行った。その結果、赤色光照射サンプルにおいて Mpphy と
MpPIF の相互作用がみられ、その相互作用は遠赤色光照射によって阻害された (図 22C)。
これらの結果から、MpPIFは活性型のMpphyと優先的に相互作用することが示唆された。
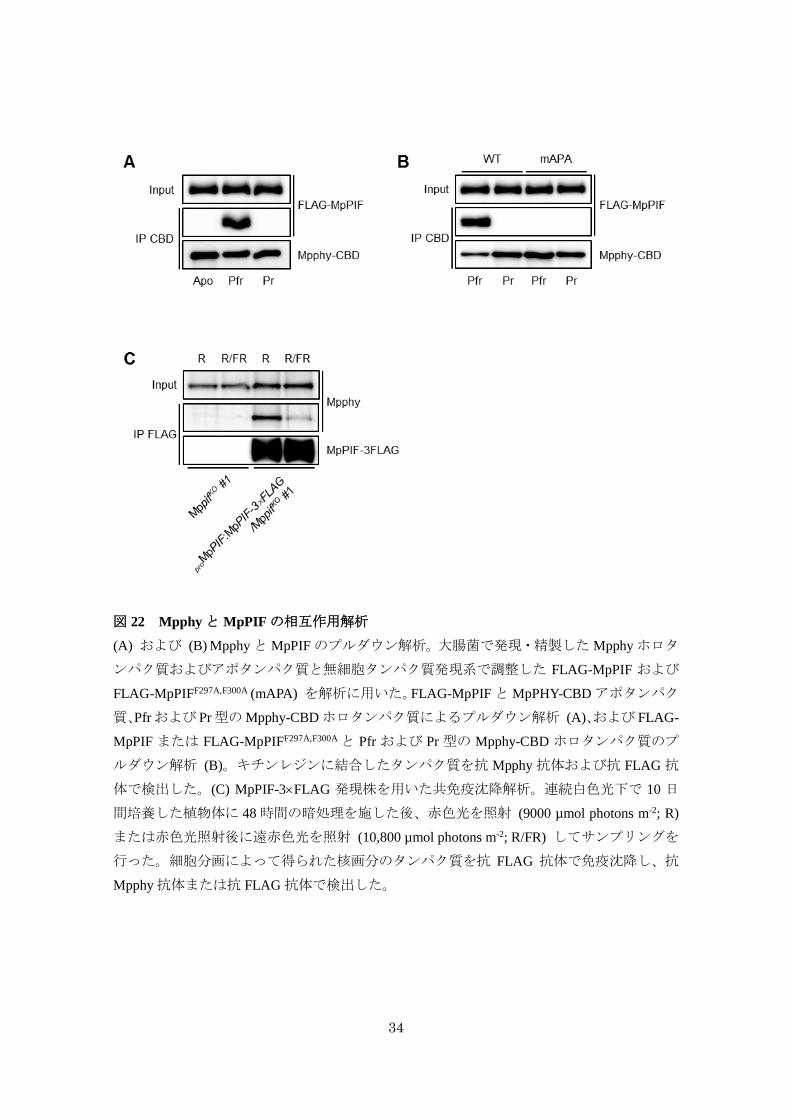
34
図 22 Mpphyと MpPIFの相互作用解析
(A) および (B) MpphyとMpPIFのプルダウン解析。大腸菌で発現・精製した Mpphyホロタ
ンパク質およびアポタンパク質と無細胞タンパク質発現系で調整した FLAG-MpPIF および
FLAG-MpPIFF297A,F300A (mAPA) を解析に用いた。FLAG-MpPIFとMpPHY-CBDアポタンパク
質、Pfrおよび Pr型のMpphy-CBDホロタンパク質によるプルダウン解析 (A)、および FLAG-
MpPIF または FLAG-MpPIFF297A,F300Aと Pfr および Pr 型の Mpphy-CBD ホロタンパク質のプ
ルダウン解析 (B)。キチンレジンに結合したタンパク質を抗 Mpphy 抗体および抗 FLAG 抗
体で検出した。(C) MpPIF-3FLAG 発現株を用いた共免疫沈降解析。連続白色光下で 10 日
間培養した植物体に 48時間の暗処理を施した後、赤色光を照射 (9000 µmol photons m-2; R)
または赤色光照射後に遠赤色光を照射 (10,800 µmol photons m-2; R/FR) してサンプリングを
行った。細胞分画によって得られた核画分のタンパク質を抗 FLAG 抗体で免疫沈降し、抗
Mpphy抗体または抗 FLAG 抗体で検出した。

35
考察
Mpphyの光に対する安定性および細胞内局在について
今回の解析から、Mpphy は被子植物のⅡ型フィトクロムと同様に光に対して安定である
ことが明らかになった (図 6)。興味深いことに、Mpphy は光に対して安定であるにも関わ
らず、被子植物のⅠ型フィトクロムにおける主要なユビキチン化部位が保存されていた
(Rattanapisit et al., 2015)。これらから、ユビキチン・プロテアソーム系を介したⅠ型フィトク
ロムの速やかな分解は、遺伝子重複によるフィトクロムの機能分化とともに陸上植物進化
の過程で獲得された可能性が考えられた。一方で、細胞内局在に関しては、Mpphyは赤色光
または遠赤色光のいずれの照射においても速やかに核内へと移行するⅠ型フィトクロム様
の性質をもつことが明らかとなった (図 7、10)。シロイヌナズナの phyA も同様に赤色光ま
たは遠赤色光のいずれの照射によっても核内へと移行するが、赤色光下では速やかに分解
されるため、遠赤色光下でのみ核内へと蓄積する (Oka et al., 2012)。これに対して、Mpphy
は赤色光下でも比較的安定なため、赤色光下でも遠赤色光下と同程度に核内に蓄積できる
可能性が考えられた。シロイヌナズナにおいて、FHY1 および FHLが phyA の遠赤色光下で
の核移行に重要であることが知られている (Hiltbrunner et al., 2006; Genoud et al., 2008;
Rausenberger et al., 2011)。近年、ヒメツリガネゴケにおける FHY1ホモログがシロイヌナズ
ナの FHY1/FHL と同様に遠赤色光照射下におけるフィトクロムの核移行に重要であること
が示された (Possart and Hiltbrunner, 2013)。FHY1 相同遺伝子はゼニゴケのゲノム上にも存在
するため、ゼニゴケの FHY1 ホモログも遠赤色光下における Mpphyの核移行に関与する可
能性が考えられた。以上の結果から、Mpphy はⅡ型フィトクロム様の光安定性をもちなが
ら、Ⅰ型フィトクロム様の細胞内局在パターンを示すというⅠ型、Ⅱ型の両方の性質を併せ
もつフィトクロムであることが示唆された。車軸藻 Mesotaenium caldariorum のフィトクロ
ムはⅠ型、Ⅱ型のどちらにも分類できない性質をもつことが報告されている (Morand et al.,
1993)。今回の結果は、陸上植物進化の初期におけるフィトクロムはⅠ型・Ⅱ型のどちらに
も分類できないという仮説を支持しており、両方の性質を併せもった祖先的な陸上植物フ
ィトクロムから維管束植物の進化の過程でⅠ型・Ⅱ型へと機能分化した可能性が考えられ
た。
Mpphyが制御する生理応答
Mpphy は無性芽の発芽や光誘導性遺伝子の発現を赤色光依存的に制御し、これらの応答
は赤色光に続く遠赤色光の照射によって阻害されたことから、Mpphy は被子植物の II 型フ
ィトクロムと同様に典型的な光可逆性を示す LFR を制御することが示された (図 11、13)。
興味深いことに、Mpphyの核移行は遠赤色光下でも観察されたにも関わらず (図 7、10)、遠
赤色光のパルス照射や連続遠赤色光は無性芽の発芽や光誘導性遺伝子の発現に有効ではな
かった (図11)。そのため、これらの応答は単純にMpphyの細胞内局在だけでは説明できず、

36
核内のフィトクロムの活性状態が重要であることが示唆された。シロイヌナズナにおいて
は、phyA は FR 照射下で核内に局在し、VLFR や FR-HIR を制御することが知られている
(Shinomura et al., 1996; Shinomura et al., 2000)。さらに、FR 照射下でのフィトクロムの核局在
や FR-HIR 様の応答はヒメツリガネゴケにおいても観察されている (Possart and Hiltbrunner,
2013)。今回の解析からは遠赤色光下で核に局在した Mpphyの生理学的な意義は明らかにで
きなかったが、過去の知見からゼニゴケの成長相転換には遠赤色光照射が必要であること
が報告されている (Chiyoda et al., 2008; Kubota et al., 2014)。そのため、遠赤色光下で核に局
在した Mpphy は成長相転換の制御に関与する可能性が考えられるが、さらなる検証が必要
である。細胞分画による細胞内局在解析の結果、遠赤色光下でのみ核内の Mpphy が何らか
の翻訳後修飾を受けることが明らかとなった (図 10)。シロイヌナズナにおいて、核内の
phyA が遠赤色光下でのみリン酸化されることが報告されており、このリン酸化は COP1 と
の相互作用を促進することが示唆されている (Saijo et al., 2008)。このことから、phyA と同
様にMpphyも遠赤色光下において核内でリン酸化される可能性が考えられた。
PIFを介した転写制御による赤色光シグナル伝達
シロイヌナズナにおいて、PIF転写因子はフィトクロムによるシグナル伝達の主要な構成
因子であることが知られている (Leivar and Monte, 2014)。しかしながら、これまでにシロイ
ヌナズナ以外の植物種による PIFの機能はほとんど報告されていない。今回、ゼニゴケに 1
分子種のみ存在する MpPIF がフィトクロムシグナル伝達に関与し、無性芽の発芽や光誘導
性遺伝子の発現を抑制することが示された (図 18)。シロイヌナズナの PIF ファミリーは暗
黒下で蓄積し、赤色光下で速やかに分解することが知られている (Leivar and Monte, 2014)。
これに対し、イネの OsPIL1 は OsphyB と相互作用せず、光に対して安定であることが報告
されている (Todaka et al., 2012)。今回、MpPIFは暗黒下で蓄積し、おそらく活性型 Mpphyと
の相互作用を介して赤色光下でのみ選択的に分解されることが示唆された (図 19、22)。こ
のことから、MpPIF はシロイヌナズナの PIF ファミリーと共通した分子機構によって活性
型フィトクロムによって機能を阻害されることが示唆された。これらの結果から、陸上植物
進化の基部に位置する苔類が、フィトクロムと PIF 転写因子の相互作用を介した転写制御
機構を既に獲得していたことが示唆された。シロイヌナズナにおいて、PIF転写因子は分解
に先立ってリン酸化されることが報告されている (Al-Sady et al., 2006; Shen et al., 2007; Shen
et al., 2008)。刺激に応じたリン酸化とそれに続く SCF 複合体による分解は多くの真核生物
において報告されている (Skaar et al., 2013)。そのため、MpPIFも分解に先立って赤色光依
存的にリン酸化される可能性が考えられるが、さらなる検証が必要である。
今回得られた知見から、ゼニゴケとシロイヌナズナにおける PIF を介したフィトクロム
シグナル伝達機構の共通点と相違点が明らかとなった (図 23)。ゼニゴケでは、赤色光下に
おいてMpphyは核内に移行し、MpPIFと APAモチーフを介して相互作用することでユビキ
チン・プロテアソーム系による MpPIFの分解を誘起する (図 19、22)。シロイヌナズナでは、

37
赤色光下において phyA および phyB の両方が核内へと移行するが、phyA は細胞質と核内の
両方で速やかに分解されるため主に phyB が核内へと蓄積する (Toledo-Ortiz et al., 2010; Oka
et al., 2012)。phyB は複数の PIF転写因子と APB モチーフを介して相互作用し、PIFのリン
酸化とそれに続く分解を誘起する (Khanna et al., 2004; Al-Sady et al., 2006; Shen et al., 2007;
Lorrain et al., 2008; Shen et al., 2008)。これらの知見から、PIF側の相互作用モチーフに違いは
あるものの、ゼニゴケとシロイヌナズナは赤色光下ではフィトクロムと PIF の光依存的な
相互作用および分解といった共通した分子機構を利用していることが示唆された。一方で、
遠赤色光のシグナル伝達はゼニゴケとシロイヌナズナで相違点がみられた。ゼニゴケでは、
Mpphy の核移行は遠赤色光下でも観察されるにもかかわらず (図 7、10)、MpPIF は遠赤色
光下では分解されず (図 19D)、無性芽の発芽や下流遺伝子の発現変動はみられなかった (図
11、13、18)。これらの知見から、遠赤色光照射下で生じる非常に少数の活性型 Mpphy は
MpPIFの分解を誘起するのに十分でないことが示唆された。シロイヌナズナでは対照的に、
遠赤色光下においては非常に少数の活性型 phyAが PIF1や PIF3の分解を誘起し (Park et al.,
2004; Al-Sady et al., 2006; Shen et al., 2008)、超低光量反応条件における種子の発芽を誘導す
ると考えられている (Oh et al., 2004; Shen et al., 2008)。今回の解析から、ゼニゴケでは遠赤
色光による超低光量反応がみられなかったことから、シロイヌナズナでは PIF の分解の高
感受性が超低光量条件下での応答を可能にすると考えられた。また、PIF4 や PIF5 は APAモ
チーフをもたないため、遠赤色光照射下では分解されず、低 R/FR 比条件下で避陰応答を制
御する (Lorrain et al., 2008)。これらの知見から、陸上植物進化の過程で 2種類のフィトクロ
ムと機能分化した複数の PIFを獲得することが様々な R/FR比に対する複雑な応答が可能に
なったと考えられた。
光によるフィトクロムの細胞内局在の制御はプラシノ藻でも観察されることから、フィ
トクロムの光依存的な核移行はストレプト植物とプラシノ藻の分岐よりも先だって獲得さ
れた形質であると推測されている (Duanmu et al., 2014)。しかしながら、ストレプト植物と
プラシノ藻のシグナル伝達機構はおそらく異なっており、プラシノ藻のフィトクロムは光
依存的に活性を示すヒスチジンキナーゼドメインがシグナル伝達に寄与していると考えら
れている (Duanmu et al., 2014)。一方で、これまで解析された全てのストレプト植物のフィ
トクロムはヒスチジンキナーゼ活性に重要なヒスチジン残基を欠損している。そのため、
PIF を介した転写制御機構は陸上植物進化の過程で新規に獲得したフィトクロムシグナル
伝達機構の 1つであり、フィトクロムシグナルの多様化に貢献した可能性が考えられた。今
回の結果から、PIFを介した転写制御による赤色光シグナル伝達機構は陸上植物進化の基部
に位置する苔類の時点で既に獲得された形質であることが示唆され、陸上植物の共通祖先
において獲得されていた可能性が考えられた。フィトクロムと PIF による赤色光シグナル
伝達機構の獲得が、植物の陸上への適応にどのように貢献したかを明らかにすることが今
後の課題である。
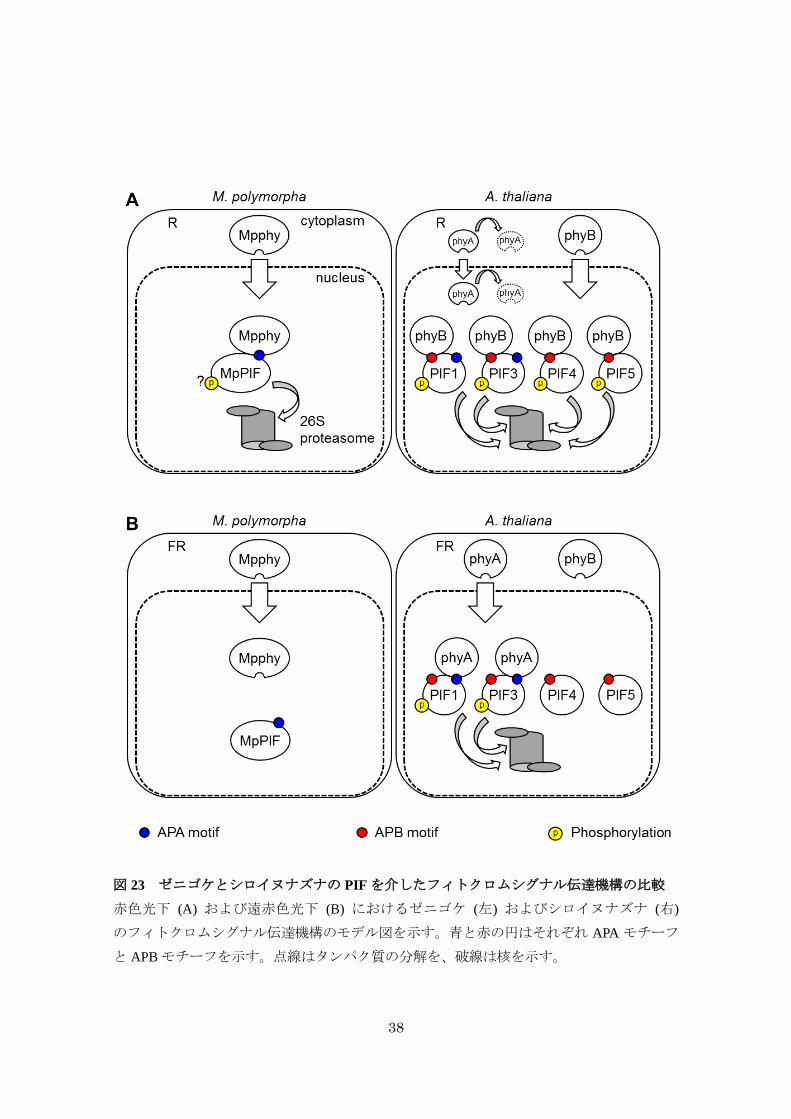
38
図 23 ゼニゴケとシロイヌナズナの PIFを介したフィトクロムシグナル伝達機構の比較
赤色光下 (A) および遠赤色光下 (B) におけるゼニゴケ (左) およびシロイヌナズナ (右)
のフィトクロムシグナル伝達機構のモデル図を示す。青と赤の円はそれぞれ APA モチーフ
と APBモチーフを示す。点線はタンパク質の分解を、破線は核を示す。

39
材料と方法
使用した植物体と培養条件
野生型雄株として Takaragaike-1 (Tak-1)、野生型雌株として Takaragaike-2 (Tak-2) を解析に
用いた (Ishizaki et al., 2008)。形質転換体の作出には Tak-1と Tak-2を掛け合わせて得られた
F1胞子を用いて行った (Ishizaki et al., 2008)。ゼニゴケの培養は 1%スクロースおよび 1.2%
寒天末を含有した 1/2Gamborg B5培地上 (Gamborg et al., 1968) で 22℃、連続白色光 (50~60
µmol photons m-2 s-1) 下で無菌的に培養した。
N末端Mpphyタンパク質による分光学的解析
MpPHY の N 末端領域 (1~612 残基) を大腸菌 ER2566 株に形質転換し、融合タンパク質
を Mukougawa et al., 2006に従って精製した。精製タンパク質溶液は試料測定用セル中で、
光源 (Intralux6000-1; Volpi AG)、赤色光フィルター (Volpi AG) および遠赤色光フィルター
(Volpi AG)を用いて、赤色光または遠赤色光を照射した。吸収スペクトルの測定には、S-
3150 Photodiode Array UV-Vis Spectrophotometer (SCINCO) を用い、操作は全て暗所において
行った。差スペクトルの測定では、遠赤色光を十分量照射後に吸収スペクトルを測定、さら
に赤色光を十分量照射した後の吸収スペクトルを測定し、両スペクトルを差し引いた値を
プロットした。このとき、それぞれの単色光を十分量照射したかを確かめるために、吸収ス
ペクトルを一定時間経過後に測定し、光異性化反応が平衡に達していることを確認した。
MpPHYのクローニング
フタバネゼニゴケのフィトクロム遺伝子 (Suzuki et al., 2001) の 3´末端領域に高い相同性
を示すゼニゴケ EST 配列、rlwb02n17 を用いて 5´RACE によって 5´末端配列を同定した。
5´RACE には、約 100 mg の葉状体から RNeasy Plant Mini Kit (QIAGEN) を用いて RNAを抽
出し、first-strand cDNA を遺伝子特異的プライマー、5´-ACCTCCATGTTCAGCT-3´、を用い
て SuperScript III (Thermo Fisher Scientific) で合成した。合成した first-strand cDNA に対して
Terminal Deoxynucleotidyl Transferase (TaKaRa) を用いて poly dC 付加を行った。poly dC を付
加した cDNAに対して遺伝子特異的プライマー、5´-GCAGGCCGAATACCTGCATG-3´、およ
び Abridged Anchor Primer (AAP; Thermo Fisher Scientific) を用いて nest PCRを行った。得ら
れた増幅断片について遺伝子特異的プライマー、5´-AGGAATTCACAGGCTGAACG-3´、およ
び Abridged Universal Amplification Primer (AUAP; Thermo Fisher Scientific) を用いて再度 nest
PCR を行い、得られた増幅断片の配列を決定した。ゲノム配列については、プライマーセッ
ト、5´-GCAGTTACGAAGGATTTTGG-3´および 5´-TTCTCTGGATGTGATCATGC-3´、を用い
て PAC ゲノムライブラリー (Okada et al., 2000) からMpPHY を含むクローンをスクリーニ
ングした。得られたクローン、pMM24-13G6、を SpeI処理して得られた約 7 kb の断片をク
ローニングし、配列を決定した。

40
使用した光源
赤色光の光源として赤色光 LED (MIL-R18、SANYO Electric、ピーク波長 657 nm、半値幅 12
nm) を用いた。遠赤色光の光源として遠赤色光 LED (MIL-IF18、SANYO Electric、ピーク波
長 734 nm、半値幅 13 nm) を用いた。光量は SKR 110 Red/Far-red Sensor (Skye instruments) を
装着した LI-250A Light Meter (LI-COR Biosciences) を用いて測定した。
系統解析
アミノ酸配列のマルチプルアライメントの作製は Geneious software (version 8.1.3;
Biomatters; http://www.geneious.com/) を用いてMUSCLE (Edgar, 2004) で行った。系統解析に
は、PIFおよび他の bHLH転写因子の bHLHドメインを用いてアライメントを作製し、PhyML
ver. 3.2.0 (Guindon et al., 2010) を用いて JTT モデルに基づき、以下の設定で作製した。Branch
Support: Bootstrap、Number of bootstraps: 1008、Number of substitution rate categories: 4、Optimize:
Topology/length/rate、Topology Search: Best。C. vulgaris および C. reinhardtii の bHLH 配列
(Pires and Dolan, 2010) を外群として用いた。解析に用いた遺伝子の accession numberは以下
の通り。 PIF1 (AEC06979), PIF3 (AEE28458), PIF4 (AEC10198), PIF5 (AEE79871), PIF6
(AEE80307), PIF7 (AED97445), PIF8 (ABG25060), SPT (ABH04569), ALC (ABI49493), bHLH23
(AEE85545), bHLH56 (AEE85546), bHLH119 (AEE85547), bHLH127 (AEE85548), and LRL1
(AEC07551), Os01g0286100 (BAF04689), Os03g0639300 (ABF97814), Os03g0782500
(ABF99196), Os05g0139100 (BAS92179), Os07g0143200 (BAF20790), Os12g0610200
(ABA99363), and Os02g0795800 (XP_015627343), Pp001G155100 (Phpat.001G155100),
Pp002G038300 (Phpat.002G038300), Pp014G091800 (Phpat.014G091800), Pp017G085300
(Phpat.017G085300), and Pp017G062100 (Phpat.017G062100)。
形質転換体の作出
・proMpEF1:MpPHY
MpPHY のコード領域を Tak-1 から抽出した RNA を鋳型に、プライマーセット 5´-
CACCATGTCGACCACCAAGGTCAC-3´および 5´-TTATTTCACACTGCCAGCAT-3´を用いて
RT-PCRによって増幅し、pENTR/D-TOPO (Thermo Fisher Scientific) にクローニングした。得
られたプラスミドを pMpGWB103 (Ishizaki et al., 2015) と LR Clonase II (Thermo Fisher
Scientific) を用いて反応させ、目的のクローンを得た。
・proMpEF1:MpPHYY241H
MpPHY のコード領域をクローニングしたプラスミドに対して、プライマーセット 5´-
CACACAAATTCCATGAGGAT-3´ お よ び 5´-CCATAACCCGGTCGTAACCT-3´ を 用 い て
Phusion Site-Directed Mutagenesis Kit (New England Biolab) を用いて変異を導入した。得ら

41
れたプラスミドを pMpGWB103 と LR Clonase II を用いて反応させ、目的のクローンを得
た。
・pro35S:MpPHY-Citrine および pro35S:MpPHYY241H-Citrine
上記で作成した MpPHY のコード領域をクローニングしたプラスミドおよび MpPHYY241H
を ク ロ ー ニ ン グ し た プ ラ ス ミ ド を 鋳 型 に 、 プ ラ イ マ ー セ ッ ト 5´-
CACCATGTCGACCACCAAGGTCAC-3´および 5´-TTTCACACTGCCAGCATCAT-3´を用いて
PCR によって増幅し、pENTR/D-TOPO にクローニングした。得られたプラスミドを
pMpGWB106 (Ishizaki et al., 2015) と LR Clonase IIを用いて反応させ、目的のクローンを
得た。
・MppifKO
Tak-1 の ゲ ノ ム DNA を 鋳 型 に 、 プ ラ イ マ ー セ ッ ト 5´-
CTAAGGTAGCGATTAATGTAGGCGTACTCGTACTC-3´/5´-
CCGGGCAAGCTTTTAATCTCTTCGAAGCTCATGTG-3´ お よ び 5´-
AACACTAGTGGCGCGAACACCTCGAAAGAGAC-3´/5´-
TTATCCCTAGGCGCGTGCAAGGAGTCTCAAGTTGG-3´を用いて MpPIF の 5´側および 3´側
の相同領域を増幅し、pJHY-TMp1 (Ishizaki et al., 2013) の PacIおよび AscIに In-Fusion HD
cloning kit (Clontech) を用いてクローニングした。得られたプラスミドをアグロバクテリウ
ムによって F1胞子に形質転換した。得られた形質転換体から、相同組み換えが生じた個体
を PCRによって選抜した (Ishizaki et al., 2013)。
・proMpPIF:MpPIF/MppifKO
MpPIF のプロモーター領域およびコード領域を Tak-1のゲノム DNAを鋳型に、プライマ
ーセット 5´-CACCTTACGAGGCAGGAGCTC-3´/5´-GGGAACGAGGTGACTCATGG-3´および
5´-TGCGAGTCTTTGGTGCTAGAG-3´/5´-TTTGGCGCGCCTGATCATATTCTGTACTTGG-3´を
用いて増幅した。増幅したプロモーター領域を pENTR/D-TOPOにクローニングし、得られ
たプラスミドの PmeI-AscIサイトに PmeIおよび AscIで処理したコード領域をクローニング
した。MpPIF のプロモーター領域およびコード領域をクローニングしたプラスミドを
pMpGWB301 (Ishizaki et al., 2015) と LR Clonase IIを用いて反応させ、目的のクローンを
得た。最終的に得られたプラスミドを MppifKO の葉状体切断片に対して形質転換した
(Kubota et al., 2013)。
・proMpPIF:MpPIF-3FLAG/MppifKO
MpPIF のプロモーター領域およびコード領域の終止コドン手前までを Tak-1 のゲノム
DNA を鋳型に、プライマーセット 5´-CACCTTACGAGGCAGGAGCTC-3´および 5´-

42
CATGCTGCTCGGAAGATGA-3´を用いて PCRによって増幅し、pENTR/D-TOPOにクローニ
ングした。得られたプラスミドを pMpGWB309 (Ishizaki et al., 2015) と LR Clonase IIを用
いて反応させ、目的のクローンを得た。最終的に得られたプラスミドをMppifKOの葉状体切
断片に対して形質転換した (Kubota et al., 2013)。
無性芽の発芽実験
緑色安全光下で無性芽を 1%スクロースおよび 1.2%寒天末を含有した 1/2Gamborg B5 培
地に播き、遠赤色光 (30 µmol photons m-2 s-1) を 15分間照射してMpphyを不活性化した後、
暗黒下で 1日間吸水させた。暗黒下で吸水した無性芽を各光条件下に移し、6日後に生長点
からの細胞分裂の有無を指標として発芽した個体数を計測した。
RNA抽出と RT-PCR
植物体を液体窒素で凍結し、Multi-Beads Shocker (安井機械) とメタルコーンを用いて 2000
rpm、1分間破砕した。RNA 抽出は TRIzol reagent (Thermo Fisher Scientific) を用いて付属の
プロトコルに従って行い、得られた RNAを RNase-Free DNase (QIAGEN) で処理した。逆転
写反応には約 1 g の RNA を使用し、ReverTra Ace (Toyobo) を用いて付属のプロトコルに
従って oligo(dT) で逆転写を行った。qRT-PCRは CFX96 Real-Time PCR Detection System (Bio-
Rad) を用いて SYBR Green I Nucleic Acid Gel Stain (Lonza) によって二本鎖 DNA 合成を定量
した。qRT-PCRの内部標準としてMpEF1およびMpAPTを用いた (Saint-Marcoux et al., 2015)。
全ての反応は 95C 30秒の後、95C 5秒、60C 30秒を 40サイクル行った。使用したプライ
マーセットを以下に示す。
MpPHY
5´-GGGCTGGGTAGGAATCAAGG-3´/5´-CCCTGAACTCAAAATGAACAACG-3´
MpPIF
5´-CAGCCGATGAGTATGGATGC-3´/5´-AGATGATGGAGCGAATGCTG-3´
LHCB
5´-GGGATTTCGTTCAGTGGATGG-3´/5´-GCTCGCTCCAATTCCGTTC-3´
POR
5´-GTCTGATGAAGCGAGCAACC-3´/5´-GACGGAACTCTAAGCGAAACC-3´
MpEF1
5´-AAGCCGTCGAAAAGAAGGAG-3´/5´-TTCAGGATCGTCCGTTATCC-3´
MpAPT
5´-CGTCGTTGTTGGAATTGAAGC-3´/5´-TTCTTGGGCTTTCGTTGAGG-3´.
全ての反応は 3 連で行い、相対発現量を Vandesompele et al., 2002 に基づいて定量した
(Vandesompele et al., 2002)。

43
MpPHY特異抗体およびMpPIF特異抗体の作製
MpPHY 抗体の抗原として、C 末端に chitin binding domain (CBD) を付加した MpPHY の
N 末端領域 (1~612 残基) を大腸菌 ER2566 株に形質転換し、融合タンパク質を Mukougawa
et al., 2006 に従って精製した。MpPIF 抗体の抗原として、N 末端に 6His タグを付加した
MpPIFの N 末端領域 (1~257) を大腸菌 Rosetta2(DE3)に形質転換した。得られた形質転換体
を 37C で対数増殖期まで培養し、1 mM の IPTG を加えて 15C で 24 時間誘導した。誘導
後の菌体を回収し、Lysis buffer (20 mM Tris-HCl pH 8.0, 150 mM NaCl, 10%(v/v) glycerol, 1 mM
DTT, 0.1 mg/mL lysozyme) に懸濁して超音波破砕した。破砕後の菌懸濁液を 16,000g、15分、
4Cで遠心後、上清を除いて沈殿を 8 M尿素含有 PBSに懸濁し、Ni-NTA agarose (QIAGEN)
を用いてアフィニティー精製した。得られた組み換えタンパク質を抗原として紀和実験動
物研究所に送付し、ウサギポリクローナル抗体を作製した。
タンパク質抽出とウエスタンブロット解析
植物体を液体窒素で凍結し、乳鉢で破砕した。粉末状の植物体と等量の抽出バッファー
(50 mM Tris-HCl pH 8.0, 150 mM NaCl, 1 mM DTT, 1 mM PMSF, 1cOmplete Protease Inhibitor
(Roche)) を加えて混和した後、16,000g、15 分、4C の遠心分離後、上清を回収した。タン
パク質溶液に等量の 2Laemmli sample buffer (100 mM Tris-HCl pH 6.8, 4%(w/v) SDS, 10%(v/v)
2-mercaptoethanol, 20%(v/v) glycerol) を加え、95Cで 5分間変性させた。Mpphyの検出には
6%の、Histone H3 の検出には 15%のアクリルアミドゲルを用いて SDS-PAGE で分離し、
PVDFメンブレン (Bio-Rad) に転写した。1次抗体には 2000倍希釈した抗 Mpphy血清また
は 5000倍希釈した anti-Histone H3 (Active Motif) を、二次抗体には 10,000倍希釈した ECL™
Anti-Rabbit IgG (GE Healthcare) を用いた。シグナルの検出には ECL Prime reagent (GE
Healthcare) を用いて化学発光を LAS4010 (FUJIFILM) により検出した。
細胞分画
寒天培地上で生育させた葉状体 (約 300 mg) を液体窒素で凍結破砕し、1 mLの Lysis buffer
(20 mM Tris-HCl pH8.0, 25%(v/v) glycerol, 10 mM MgCl2, 250 mM sucrose, 5 mM DTT, 1 mM
PMSF, 20 M MG132, 1cOmplete EDTA-free Protease Inhibitor) に懸濁した。懸濁液を 70 m
cell strainer に通し、フロースルーを 1500g、10 分、4C で遠心分離した。得られた上清を
「細胞質画分」として回収し、等量の 2Laemmli sample bufferを加え、95Cで 5分間変性
させた。得られた沈殿は 1 mLのWash buffer (20 mM Tris-HCl pH8.0, 25%(v/v) glycerol, 2.5
mM MgCl2, 0.2%(v/v) Triton X-100, 5 mM DTT, 1mM PMSF, 20 M MG132, 1cOmplete EDTA-
free Protease Inhibitor) で 2回洗浄し、50 Lの Extraction buffer (20 mM Tris-HCl pH8.0, 5 mM
EDTA,150 mM NaCl, 1%(v/v) Triton X-100, 0.5%(w/v) sodium deoxycholate, 0.1%(w/v) SDS, 5
mM DTT, 1mM PMSF, 20 M MG132, 1cOmplete Protease Inhibitor) に懸濁して氷上で 10分
間静置した。得られた溶液を「核画分」とし、等量の 2Laemmli sample bufferを加え、95C
で 5 分間変性させた。それぞれの画分を 16,000g、15 分、室温で遠心し、上清をウエスタ

44
ンブロット解析に用いた。1 次抗体には 2000 倍希釈した抗 Mpphy血清、2000 倍希釈した
抗MpPIF血清または 5000倍希釈した anti-Histone H3 (Active Motif) を、二次抗体には 10,000
倍希釈した ECL™ Anti-Rabbit IgG (GE Healthcare) を用いた。
pull-down解析
baitタンパク質として、C末端に CBD を融合した N 末端MpPHY(1-612) アポタンパク質
および発色団として PBを結合させたホロタンパク質を大腸菌で発現・精製した。組み換
えタンパク質は chitin resin (New England Biolabs) と 4C で 1時間結合させ、Binding buffer
(20 mM Tris-HCl pH 8.0, 150 mM NaCl, 0.1%(v/v) Triton X-100, 0.01%(w/v) BSA) で 3回洗浄し
た。アフィニティー結合した Mpphy タンパク質の濃度はウエスタンブロット解析によって
推定した。preyタンパク質として、N 末端に FLAG を融合したMpPIFおよび MpPIFF297A,F300A
を無細胞タンパク質発現系 (BioSieg) で調整した。反応溶液を 15,000g、 10 分、4C で遠心
し、ウエスタンブロット解析で濃度を推定した後、pull-down 解析に用いた。Mpphy(N612)-
CBDホロタンパク質は解析前に赤色光 (9000 mol photons m-2; Pfr) または赤色光に続いて
遠赤色光 (10,800 mol photons m-2; Pr) をパルス照射して用いた。10 LのMpphy(N612)-CBD
アポタンパク質またはホロタンパク質を、10 L の FLAG-MpPIF または MpPIFF297A,F300Aと
混合し、80 Lの Binding bufferを加えて 4C、暗黒下で 1時間静置した。レジンを 1 mLの
Binding bufferで 3回洗浄し、10 Lの 2Laemmli sample buffer を加え、95C で 5分間変性
させ、ウエスタンブロット解析に用いた。
共免疫沈降
寒天培地上で生育させた葉状体 (約 2.0 g) を液体窒素で凍結破砕し、上記の細胞分画を行
った。得られた核の沈殿を 1 mLの Extraction buffer (20 mM Tris-HCl pH8.0, 5 mM EDTA,150
mM NaCl, 1%(v/v) Triton X-100, 0.5%(w/v) sodium deoxycholate, 5 mM DTT, 1mM PMSF, 50 M
MG132, 1cOmplete Protease Inhibitor) に懸濁し、氷上で 10分間静置した。懸濁液を 16,000g、
10 分、4Cで遠心し、上清を免疫沈降に用いた。共免疫沈降はMACS DYKDDDDK Isolation
Kit (Miltenyi Biotec) を用いて付属のプロトコルに従って行った。
蛍光顕微鏡観察
無性芽を連続白色光下で 1日間培養後、6日間の暗処理を施し、赤色光または遠赤色光を
10 分間照射した。GaAsP 検出器を取り付けた Olympus FV1000 共焦点レーザー顕微鏡を用
いて、515 nmのレーザーで励起した Citrine由来の蛍光を検出した。

45
文献
Al-Sady, B., Ni, W., Kircher, S., Schafer, E., and Quail, P.H. (2006). Photoactivated phytochrome induces
rapid PIF3 phosphorylation prior to proteasome-mediated degradation. Mol. Cell 23: 439-446.
Alosi, M.C., and Neale, D.B. (1992). Light- and phytochrome-mediated gene expression in Douglas-fir
seedlings. Physiol. Plant. 86: 71-76.
Bae, G., and Choi, G. (2008). Decoding of light signals by plant phytochromes and their interacting
proteins. Annu. Rev. Plant Biol. 59: 281-311.
Casal, J.J., Sánchez, R.A., and Botto, J.F. (1998). Modes of action of phytochromes. J. Exp. Bot. 49:
127-138.
Chen, M., Tao, Y., Lim, J., Shaw, A., and Chory, J. (2005). Regulation of phytochrome B nuclear
localization through light-dependent unmasking of nuclear-localization signals. Curr. Biol. 15:
637-642.
Chiyoda, S., Ishizaki, K., Kataoka, H., Yamato, K.T., and Kohchi, T. (2008). Direct transformation of
the liverwort Marchantia polymorpha L. by particle bombardment using immature thalli
developing from spores. Plant Cell Rep. 27: 1467-1473.
Christensen, S., LaVerne, E., Boyd, G., and Silverthorne, J. (2002). Ginkgo biloba retains functions of
both type I and type II flowering plant phytochrome. Plant Cell Physiol. 43: 768-777.
Christie, J.M., Blackwood, L., Petersen, J., and Sullivan, S. (2015). Plant flavoprotein photoreceptors.
Plant Cell Physiol. 56: 401-413.
Clack, T., Mathews, S., and Sharrock, R.A. (1994). The phytochrome apoprotein family in Arabidopsis
is encoded by five genes: the sequences and expression of PHYD and PHYE. Plant Mol. Biol. 25:
413-427.
De Greef, J., Butler, W.L., and Roth, T.F. (1971). Control of senescence in Marchantia by phytochrome.
Plant Physiol. 48: 407-412.
Duanmu, D., Bachy, C., Sudek, S., Wong, C.H., Jimenez, V., Rockwell, N.C., Martin, S.S., Ngan, C.Y.,
Reistetter, E.N., van Baren, M.J., Price, D.C., Wei, C.L., Reyes-Prieto, A., Lagarias, J.C., and
Worden, A.Z. (2014). Marine algae and land plants share conserved phytochrome signaling
systems. Proc. Natl. Acad. Sci. USA 111: 15827-15832.
Edgar, R.C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput.
Nucleic Acids Res. 32: 1792-1797.
Fankhauser, C., and Chen, M. (2008). Transposing phytochrome into the nucleus. Trends Plant Sci. 13:
596-601.
Fredericq, H., and De Greef, J. (1966). Red (R), far-red (FR) photoreversible control of growth and
chlorophyll content in light-grown thalli of Marchantia polymorpha L. Naturwissenschaften 53:
337.

46
Gamborg, O.L., Miller, R.A., and Ojima, K. (1968). Nutrient requirements of suspension cultures of
soybean root cells. Exp. Cell Res. 50: 151-158.
Genoud, T., Schweizer, F., Tscheuschler, A., Debrieux, D., Casal, J.J., Schafer, E., Hiltbrunner, A.,
and Fankhauser, C. (2008). FHY1 mediates nuclear import of the light-activated phytochrome A
photoreceptor. PLoS Genet. 4: e1000143.
Gil, P., Kircher, S., Adam, E., Bury, E., Kozma-Bognar, L., Schafer, E., and Nagy, F. (2000).
Photocontrol of subcellular partitioning of phytochrome-B:GFP fusion protein in tobacco
seedlings. Plant J. 22: 135-145.
Guindon, S., Dufayard, J.F., Lefort, V., Anisimova, M., Hordijk, W., and Gascuel, O. (2010). New
algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance
of PhyML 3.0. Syst. Biol. 59: 307-321.
Hartmann, E., and Jenkins, G.I. (1984). Photomorphogenesis of Mosses and Liverworts. In The
Experimental Biology of Bryophytes, A. F. Dyer and J. G. Duckett, eds. (Academic Press, London),
pp. 203-228.
Hiltbrunner, A., Tscheuschler, A., Viczian, A., Kunkel, T., Kircher, S., and Schafer, E. (2006). FHY1
and FHL act together to mediate nuclear accumulation of the phytochrome A photoreceptor. Plant
Cell Physiol. 47: 1023-1034.
Huang, X., Ouyang, X., and Deng, X.W. (2014). Beyond repression of photomorphogenesis: role
switching of COP/DET/FUS in light signaling. Curr. Opin. Plant Biol. 21: 96-103.
Huq, E., and Quail, P.H. (2002). PIF4, a phytochrome-interacting bHLH factor, functions as a negative
regulator of phytochrome B signaling in Arabidopsis. EMBO J. 21: 2441-2450.
Huq, E., Al-Sady, B., and Quail, P.H. (2003). Nuclear translocation of the photoreceptor phytochrome B
is necessary for its biological function in seedling photomorphogenesis. Plant J. 35: 660-664.
Huq, E., Al-Sady, B., Hudson, M., Kim, C., Apel, K., and Quail, P.H. (2004). Phytochrome-interacting
factor 1 is a critical bHLH regulator of chlorophyll biosynthesis. Science 305: 1937-1941.
Ishizaki, K., Chiyoda, S., Yamato, K.T., and Kohchi, T. (2008). Agrobacterium-mediated transformation
of the haploid liverwort Marchantia polymorpha L., an emerging model for plant biology. Plant
Cell Physiol. 49: 1084-1091.
Ishizaki, K., Johzuka-Hisatomi, Y., Ishida, S., Iida, S., and Kohchi, T. (2013). Homologous
recombination-mediated gene targeting in the liverwort Marchantia polymorpha L. Sci. Rep. 3:
1532.
Ishizaki, K., Nishihama, R., Ueda, M., Inoue, K., Ishida, S., Nishimura, Y., Shikanai, T., and Kohchi,
T. (2015). Development of gateway binary vector series with four different selection markers for
the liverwort Marchantia polymorpha. PLoS ONE 10: e0138876.
Ito, S., Song, Y.H., and Imaizumi, T. (2012). LOV domain-containing F-box proteins: light-dependent
protein degradation modules in Arabidopsis. Mol. Plant 5: 573-582.

47
Jaedicke, K., Lichtenthaler, A.L., Meyberg, R., Zeidler, M., and Hughes, J. (2012). A phytochrome-
phototropin light signaling complex at the plasma membrane. Proc. Natl. Acad. Sci. USA 109:
12231-12236.
Jenkins, G.I. (2014). The UV-B photoreceptor UVR8: from structure to physiology. Plant Cell 26: 21-37.
Kami, C., Lorrain, S., Hornitschek, P., and Fankhauser, C. (2010). Light-regulated plant growth and
development. Curr. Top. Dev. Biol. 91: 29-66.
Karas, B., Amyot, L., Johansen, C., Sato, S., Tabata, S., Kawaguchi, M., and Szczyglowski, K. (2009).
Conservation of lotus and Arabidopsis basic helix-loop-helix proteins reveals new players in root
hair development. Plant Physiol. 151: 1175-1185.
Kebrom, T.H., Burson, B.L., and Finlayson, S.A. (2006). Phytochrome B represses Teosinte Branched1
expression and induces sorghum axillary bud outgrowth in response to light signals. Plant Physiol.
140: 1109-1117.
Kebrom, T.H., Brutnell, T.P., and Finlayson, S.A. (2010). Suppression of sorghum axillary bud
outgrowth by shade, phyB and defoliation signalling pathways. Plant Cell Environ. 33: 48-58.
Khanna, R., Huq, E., Kikis, E.A., Al-Sady, B., Lanzatella, C., and Quail, P.H. (2004). A novel
molecular recognition motif necessary for targeting photoactivated phytochrome signaling to
specific basic helix-loop-helix transcription factors. Plant Cell 16: 3033-3044.
Kim, L., Kircher, S., Toth, R., Adam, E., Schafer, E., and Nagy, F. (2000). Light-induced nuclear import
of phytochrome-A:GFP fusion proteins is differentially regulated in transgenic tobacco and
Arabidopsis. Plant J. 22: 125-133.
Kircher, S., Kozma-Bognar, L., Kim, L., Adam, E., Harter, K., Schafer, E., and Nagy, F. (1999). Light
quality-dependent nuclear import of the plant photoreceptors phytochrome A and B. Plant Cell 11:
1445-1456.
Klose, C., Viczian, A., Kircher, S., Schafer, E., and Nagy, F. (2015). Molecular mechanisms for mediating
light-dependent nucleo/cytoplasmic partitioning of phytochrome photoreceptors. New Phytol.
206: 965-971.
Kohchi, T., Mukougawa, K., Frankenberg, N., Masuda, M., Yokota, A., and Lagarias, J.C. (2001).
The Arabidopsis HY2 gene encodes phytochromobilin synthase, a ferredoxin-dependent biliverdin
reductase. Plant Cell 13: 425-436.
Kubota, A., Ishizaki, K., Hosaka, M., and Kohchi, T. (2013). Efficient Agrobacterium-mediated
transformation of the liverwort Marchantia polymorpha using regenerating thalli. Biosci.
Biotechnol. Biochem. 77: 167-172.
Kubota, A., Kita, S., Ishizaki, K., Nishihama, R., Yamato, K.T., and Kohchi, T. (2014). Co-option of a
photoperiodic growth-phase transition system during land plant evolution. Nat. Commun. 5: 3668.
Lagarias, J.C., and Lagarias, D.M. (1989). Self-assembly of synthetic phytochrome holoprotein in vitro.
Proc. Natl. Acad. Sci. USA 86: 5778-5780.

48
Leivar, P., and Quail, P.H. (2011). PIFs: pivotal components in a cellular signaling hub. Trends Plant Sci.
16: 19-28.
Leivar, P., and Monte, E. (2014). PIFs: systems integrators in plant development. Plant Cell 26: 56-78.
Leivar, P., Monte, E., Al-Sady, B., Carle, C., Storer, A., Alonso, J.M., Ecker, J.R., and Quail, P.H.
(2008). The Arabidopsis phytochrome-interacting factor PIF7, together with PIF3 and PIF4,
regulates responses to prolonged red light by modulating phyB levels. Plant Cell 20: 337-352.
Li, F.W., Melkonian, M., Rothfels, C.J., Villarreal, J.C., Stevenson, D.W., Graham, S.W., Wong, G.K.,
Pryer, K.M., and Mathews, S. (2015). Phytochrome diversity in green plants and the origin of
canonical plant phytochromes. Nat. Commun. 6: 7852.
Lorrain, S., Allen, T., Duek, P.D., Whitelam, G.C., and Fankhauser, C. (2008). Phytochrome-mediated
inhibition of shade avoidance involves degradation of growth-promoting bHLH transcription
factors. Plant J. 53: 312-323.
Lu, X.D., Zhou, C.M., Xu, P.B., Luo, Q., Lian, H.L., and Yang, H.Q. (2015). Red-light-dependent
interaction of phyB with SPA1 promotes COP1-SPA1 dissociation and photomorphogenic
development in Arabidopsis. Mol. Plant 8: 467-478.
Mancinelli, A.L. (1994). The physiology of phytochrome action. In Photomorphogenesis in Plants, 2nd
ed., R.E. Kendrick and G.H.M. Kronenberg, eds. (Dordrecht, The Netherlands: Kluwer Academic
Publishers), pp. 211–269.
Mateos, J.L., Luppi, J.P., Ogorodnikova, O.B., Sineshchekov, V.A., Yanovsky, M.J., Braslavsky, S.E.,
Gartner, W., and Casal, J.J. (2006). Functional and biochemical analysis of the N-terminal
domain of phytochrome A. J. Biol. Chem. 281: 34421-34429.
Matsushita, T., Mochizuki, N., and Nagatani, A. (2003). Dimers of the N-terminal domain of
phytochrome B are functional in the nucleus. Nature 424: 571-574.
Mittmann, F., Dienstbach, S., Weisert, A., and Forreiter, C. (2009). Analysis of the phytochrome gene
family in Ceratodon purpureus by gene targeting reveals the primary phytochrome responsible for
photo- and polarotropism. Planta 230: 27-37.
Mittmann, F., Brucker, G., Zeidler, M., Repp, A., Abts, T., Hartmann, E., and Hughes, J. (2004).
Targeted knockout in Physcomitrella reveals direct actions of phytochrome in the cytoplasm. Proc.
Natl. Acad. Sci. USA 101: 13939-13944.
Morand, L.Z., Kidd, D.G., and Lagarias, J.C. (1993). Phytochrome levels in the green alga Mesotaenium
caldariorum are light regulated. Plant Physiol. 101: 97-104.
Muller, R., Fernandez, A.P., Hiltbrunner, A., Schafer, E., and Kretsch, T. (2009). The histidine kinase-
related domain of Arabidopsis phytochrome a controls the spectral sensitivity and the subcellular
distribution of the photoreceptor. Plant Physiol. 150: 1297-1309.
Muramoto, T., Kohchi, T., Yokota, A., Hwang, I., and Goodman, H.M. (1999). The Arabidopsis
photomorphogenic mutant hy1 is deficient in phytochrome chromophore biosynthesis as a result

49
of a mutation in a plastid heme oxygenase. Plant Cell 11: 335-348.
Nagatani, A. (2004). Light-regulated nuclear localization of phytochromes. Curr. Opin. Plant Biol. 7: 708-
711.
Nagatani, A. (2010). Phytochrome: structural basis for its functions. Curr. Opin. Plant Biol. 13: 565-570.
Nagatani, A., Reed, J.W., and Chory, J. (1993). Isolation and initial characterization of arabidopsis
mutants that are deficient in phytochrome A. Plant Physiol. 102: 269-277.
Nakamura, Y., Kato, T., Yamashino, T., Murakami, M., and Mizuno, T. (2007). Characterization of a
set of phytochrome-interacting factor-like bHLH proteins in Oryza sativa. Biosci. Biotechnol.
Biochem. 71: 1183-1191.
Ni, M., Tepperman, J.M., and Quail, P.H. (1998). PIF3, a phytochrome-interacting factor necessary for
normal photoinduced signal transduction, is a novel basic helix-loop-helix protein. Cell 95: 657-
667.
Ni, M., Tepperman, J.M., and Quail, P.H. (1999). Binding of phytochrome B to its nuclear signalling
partner PIF3 is reversibly induced by light. Nature 400: 781-784.
Ninnemann, H., and Halbsguth, W. (1965). Rolle des Phytochroms beim Etiolement von Marchantia
polymorpha. Naturwissenschaften 52: 110-111.
Nishihama, R., Ishizaki, K., Hosaka, M., Matsuda, Y., Kubota, A., and Kohchi, T. (2015).
Phytochrome-mediated regulation of cell division and growth during regeneration and sporeling
development in the liverwort Marchantia polymorpha. J. Plant Res. 128: 407-421.
Oh, E., Kim, J., Park, E., Kim, J.I., Kang, C., and Choi, G. (2004). PIL5, a phytochrome-interacting
basic helix-loop-helix protein, is a key negative regulator of seed germination in Arabidopsis
thaliana. Plant Cell 16: 3045-3058.
Oka, Y., Kong, S.G., and Matsushita, T. (2011). A non-covalently attached chromophore can mediate
phytochrome B signaling in Arabidopsis. Plant Cell Physiol. 52: 2088-2102.
Oka, Y., Matsushita, T., Mochizuki, N., Suzuki, T., Tokutomi, S., and Nagatani, A. (2004). Functional
analysis of a 450-amino acid N-terminal fragment of phytochrome B in Arabidopsis. Plant Cell
16: 2104-2116.
Oka, Y., Ono, Y., Toledo-Ortiz, G., Kokaji, K., Matsui, M., Mochizuki, N., and Nagatani, A. (2012).
Arabidopsis phytochrome a is modularly structured to integrate the multiple features that are
required for a highly sensitized phytochrome. Plant Cell 24: 2949-2962.
Okada, S., Fujisawa, M., Sone, T., Nakayama, S., Nishiyama, R., Takenaka, M., Yamaoka, S., Sakaida,
M., Kono, K., Takahama, M., Yamato, K.T., Fukuzawa, H., Brennicke, A., and Ohyama, K.
(2000). Construction of male and female PAC genomic libraries suitable for identification of Y-
chromosome-specific clones from the liverwort, Marchantia polymorpha. Plant J. 24: 421-428.
Otto, K.-R., and Halbsguth, W. (1976). Die Förderung der Bildung von Primärrhizoiden an Brutkörpern
von Marchantia polymorpha L. durch Licht und IES. Zeitschrift für Pflanzenphysiologie 80: 197-

50
205.
Park, E., Kim, J., Lee, Y., Shin, J., Oh, E., Chung, W.I., Liu, J.R., and Choi, G. (2004). Degradation of
phytochrome interacting factor 3 in phytochrome-mediated light signaling. Plant Cell Physiol. 45:
968-975.
Parks, B.M., and Quail, P.H. (1993). hy8, a new class of arabidopsis long hypocotyl mutants deficient in
functional phytochrome A. Plant Cell 5: 39-48.
Pasentsis, K., Paulo, N., Algarra, P., Dittrich, P., and Thummler, F. (1998). Characterization and
expression of the phytochrome gene family in the moss Ceratodon purpureus. Plant J. 13: 51-61.
Pfeiffer, A., Nagel, M.K., Popp, C., Wust, F., Bindics, J., Viczian, A., Hiltbrunner, A., Nagy, F., Kunkel,
T., and Schafer, E. (2012). Interaction with plant transcription factors can mediate nuclear import
of phytochrome B. Proc. Natl. Acad. Sci. USA 109: 5892-5897.
Pires, N., and Dolan, L. (2010). Origin and diversification of basic-helix-loop-helix proteins in plants.
Mol. Biol. Evol. 27: 862-874.
Possart, A., and Hiltbrunner, A. (2013). An evolutionarily conserved signaling mechanism mediates far-
red light responses in land plants. Plant Cell 25: 102-114.
Qiu, Y.L., Li, L., Wang, B., Chen, Z., Knoop, V., Groth-Malonek, M., Dombrovska, O., Lee, J., Kent,
L., Rest, J., Estabrook, G.F., Hendry, T.A., Taylor, D.W., Testa, C.M., Ambros, M., Crandall-
Stotler, B., Duff, R.J., Stech, M., Frey, W., Quandt, D., and Davis, C.C. (2006). The deepest
divergences in land plants inferred from phylogenomic evidence. Proc. Natl. Acad. Sci. USA 103:
15511-15516.
Quail, P.H. (2010). Phytochromes. Curr. Biol. 20: R504-507.
Rattanapisit, K., Cho, M.H., and Bhoo, S.H. (2015). Lysine 206 in Arabidopsis phytochrome A is the
major site for ubiquitin-dependent protein degradation. J. Biochem.
Rausenberger, J., Tscheuschler, A., Nordmeier, W., Wust, F., Timmer, J., Schafer, E., Fleck, C., and
Hiltbrunner, A. (2011). Photoconversion and nuclear trafficking cycles determine phytochrome
A's response profile to far-red light. Cell 146: 813-825.
Reddy, R.K., and Sharma, R. (1998). Spatial distribution and temporal regulation of phytochromes A and
B levels in maize seedlings. Plant Physiol. Biochem. 36: 737–745.
Rosler, J., Jaedicke, K., and Zeidler, M. (2010). Cytoplasmic phytochrome action. Plant Cell Physiol. 51:
1248-1254.
Saijo, Y., Zhu, D., Li, J., Rubio, V., Zhou, Z., Shen, Y., Hoecker, U., Wang, H., and Deng, X.W. (2008).
Arabidopsis COP1/SPA1 complex and FHY1/FHY3 associate with distinct phosphorylated forms
of phytochrome A in balancing light signaling. Mol. Cell 31: 607-613.
Saint-Marcoux, D., Proust, H., Dolan, L., and Langdale, J.A. (2015). Identification of reference genes
for real-time quantitative PCR experiments in the liverwort Marchantia polymorpha. PLoS ONE
10.

51
Sawers, R.J., Linley, P.J., Farmer, P.R., Hanley, N.P., Costich, D.E., Terry, M.J., and Brutnell, T.P.
(2002). Elongated mesocotyl1, a phytochrome-deficient mutant of maize. Plant Physiol. 130: 155-
163.
Sharrock, R.A., and Clack, T. (2002). Patterns of expression and normalized levels of the five Arabidopsis
phytochromes. Plant Physiol. 130: 442-456.
Sheerin, D.J., Menon, C., zur Oven-Krockhaus, S., Enderle, B., Zhu, L., Johnen, P., Schleifenbaum,
F., Stierhof, Y.D., Huq, E., and Hiltbrunner, A. (2015). Light-activated phytochrome A and B
interact with members of the SPA family to promote photomorphogenesis in Arabidopsis by
reorganizing the COP1/SPA complex. Plant Cell 27: 189-201.
Shen, H., Zhu, L., Castillon, A., Majee, M., Downie, B., and Huq, E. (2008). Light-induced
phosphorylation and degradation of the negative regulator PHYTOCHROME-INTERACTING
FACTOR1 from Arabidopsis depend upon its direct physical interactions with photoactivated
phytochromes. Plant Cell 20: 1586-1602.
Shen, Y., Khanna, R., Carle, C.M., and Quail, P.H. (2007). Phytochrome induces rapid PIF5
phosphorylation and degradation in response to red-light activation. Plant Physiol. 145: 1043-1051.
Shimizu-Sato, S., Huq, E., Tepperman, J.M., and Quail, P.H. (2002). A light-switchable gene promoter
system. Nat. Biotechnol. 20: 1041-1044.
Shinomura, T., Uchida, K., and Furuya, M. (2000). Elementary processes of photoperception by
phytochrome A for high-irradiance response of hypocotyl elongation in Arabidopsis. Plant Physiol.
122: 147-156.
Shinomura, T., Nagatani, A., Hanzawa, H., Kubota, M., Watanabe, M., and Furuya, M. (1996). Action
spectra for phytochrome A- and B-specific photoinduction of seed germination in Arabidopsis
thaliana. Proc. Natl. Acad. Sci. USA 93: 8129-8133.
Skaar, J.R., Pagan, J.K., and Pagano, M. (2013). Mechanisms and function of substrate recruitment by
F-box proteins. Nat. Rev. Mol. Cell Biol. 14: 369-381.
Su, Y.S., and Lagarias, J.C. (2007). Light-independent phytochrome signaling mediated by dominant
GAF domain tyrosine mutants of Arabidopsis phytochromes in transgenic plants. Plant Cell 19:
2124-2139.
Suetsugu, N., and Wada, M. (2007). Phytochrome-dependent photomovement responses mediated by
phototropin family proteins in cryptogam plants. Photochem. Photobiol. 83: 87-93.
Sugano, S.S., Shirakawa, M., Takagi, J., Matsuda, Y., Shimada, T., Hara-Nishimura, I., and Kohchi,
T. (2014). CRISPR/Cas9-mediated targeted mutagenesis in the liverwort Marchantia polymorpha
L. Plant Cell Physiol. 55: 475-481.
Suzuki, T., Takio, S., Yamamoto, I., and Satoh, T. (2001). Characterization of cDNA of the liverwort
phytochrome gene, and phytochrome involvement in the light-dependent and light-independent
protochlorophyllide oxidoreductase gene expression in Marchantia paleacea var. diptera. Plant

52
Cell Physiol. 42: 576-582.
Takano, M., Inagaki, N., Xie, X., Kiyota, S., Baba-Kasai, A., Tanabata, T., and Shinomura, T. (2009).
Phytochromes are the sole photoreceptors for perceiving red/far-red light in rice. Proc. Natl. Acad.
Sci. USA 106: 14705-14710.
Takano, M., Inagaki, N., Xie, X., Yuzurihara, N., Hihara, F., Ishizuka, T., Yano, M., Nishimura, M.,
Miyao, A., Hirochika, H., and Shinomura, T. (2005). Distinct and cooperative functions of
phytochromes A, B, and C in the control of deetiolation and flowering in rice. Plant Cell 17: 3311-
3325.
Thummler, F., Dufner, M., Kreisl, P., and Dittrich, P. (1992). Molecular cloning of a novel phytochrome
gene of the moss Ceratodon purpureus which encodes a putative light-regulated protein kinase.
Plant Mol. Biol. 20: 1003-1017.
Todaka, D., Nakashima, K., Maruyama, K., Kidokoro, S., Osakabe, Y., Ito, Y., Matsukura, S., Fujita,
Y., Yoshiwara, K., Ohme-Takagi, M., Kojima, M., Sakakibara, H., Shinozaki, K., and
Yamaguchi-Shinozaki, K. (2012). Rice phytochrome-interacting factor-like protein OsPIL1
functions as a key regulator of internode elongation and induces a morphological response to
drought stress. Proc. Natl. Acad. Sci. USA 109: 15947-15952.
Toledo-Ortiz, G., Kiryu, Y., Kobayashi, J., Oka, Y., Kim, Y., Nam, H.G., Mochizuki, N., and Nagatani,
A. (2010). Subcellular sites of the signal transduction and degradation of phytochrome A. Plant
Cell Physiol. 51: 1648-1660.
Tsuboi, H., Nakamura, S., Schafer, E., and Wada, M. (2012). Red light-induced phytochrome relocation
into the nucleus in Adiantum capillus-veneris. Mol. Plant 5: 611-618.
Uenaka, H., and Kadota, A. (2007). Functional analyses of the Physcomitrella patens phytochromes in
regulating chloroplast avoidance movement. Plant J. 51: 1050-1061.
Uenaka, H., Wada, M., and Kadota, A. (2005). Four distinct photoreceptors contribute to light-induced
side branch formation in the moss Physcomitrella patens. Planta 222: 623-631.
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., and Speleman, F.
(2002). Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of
multiple internal control genes. Genome Biol. 3.
Whitelam, G.C., Johnson, E., Peng, J., Carol, P., Anderson, M.L., Cowl, J.S., and Harberd, N.P.
(1993). Phytochrome A null mutants of Arabidopsis display a wild-type phenotype in white light.
Plant Cell 5: 757-768.
Xie, X., Kagawa, T., and Takano, M. (2014). The phytochrome B/phytochrome C heterodimer is
necessary for phytochrome C-mediated responses in rice seedlings. PLoS ONE 9: e97264.
Yamaguchi, R., Nakamura, M., Mochizuki, N., Kay, S.A., and Nagatani, A. (1999). Light-dependent
translocation of a phytochrome B-GFP fusion protein to the nucleus in transgenic Arabidopsis. J.
Cell Biol. 145: 437-445.

53
Yamashino, T., Matsushika, A., Fujimori, T., Sato, S., Kato, T., Tabata, S., and Mizuno, T. (2003). A
Link between circadian-controlled bHLH factors and the APRR1/TOC1 quintet in Arabidopsis
thaliana. Plant Cell Physiol. 44: 619-629.
Zhu, Y., Tepperman, J.M., Fairchild, C.D., and Quail, P.H. (2000). Phytochrome B binds with greater
apparent affinity than phytochrome A to the basic helix-loop-helix factor PIF3 in a reaction
requiring the PAS domain of PIF3. Proc. Natl. Acad. Sci. USA 97: 13419-13424.

54
謝辞
本研究は京都大学大学院生命科学研究科遺伝子特性学分野の河内孝之教授の下で行われま
した。河内教授には長年にわたり多くのご指導・ご支援を賜りました。心より御礼申し上げ
ます。本研究を遂行するにあたり、石崎公庸准教授(神戸大学大学院理学研究科)、西浜竜
一講師には論文執筆について多くのご指導とご提言をいただきました。心より感謝いたし
ます。山岡尚平助教、末次憲之特定助教にはセミナーやミーティングの場を通じて多くのご
助言をいただきました。また福澤秀哉教授(京都大学生命科学研究科)、大和勝幸准教授(近
畿大学生物理工学部)には当研究室にご在籍中に様々なご助言をいただきました。深く御礼
申し上げます。荒木崇教授(京都大学生命科学研究科)、神戸大朋准教授(京都大学生命科
学研究科)には副指導教員としてご指導いただきました。深く御礼申し上げます。本研究を
行うにあたり、植物用バイナリ―ベクターを中川強教授(島根大学)から分与していただき
ました。この場を借りて御礼申し上げます。また本研究は日本学術振興会より特別研究員と
しての支援をいただきました。お礼申し上げます。
本研究が行われました遺伝子特性学分野の皆様には、研究生活を通じて多くのご助言、ご協
力をいただきました。多大な支援を頂いた全ての皆様のおかげで楽しく研究生活を送るこ
とができました。深く感謝いたします。
本学位論文は以下の学術論文の内容に基づいて書かれたものである。
Keisuke Inoue, Ryuichi Nishihama, Hideo Kataoka, Masashi Hosaka, Ryo Manabe, Mika Nomoto,
Yasuomi Tada, Kimitsune Ishizaki, and Takayuki Kohchi
Phytochrome Signaling Is Mediated by PHYTOCHROME INTERACTING FACTOR in the
Liverwort Marchantia polymorpha
The Plant Cell, Vol. 28: 1406-1421, June 2016